Accueil > 02 - Livre Deux : SCIENCES > Développement et génétique > Comment le Vivant mesure le Temps
Comment le Vivant mesure le Temps
samedi 12 mai 2018, par









Comment le Vivant mesure le Temps
Avertissement : En soulignant l’existence d’horloges biologiques, nous ne voulons pas dire que la vie « crée » le temps mais seulement qu’elle le mesure ou le bat. Pas plus qu’il n’était exact que ce serait la conscience humaine qui produirait le temps comme le croyait Bergson, ce n’est pas le vivant qui fabrique le rythme et l’écoulement (la flèche) du temps. Le temps est marqué par toutes les rétroactions emboîtées, depuis celles de groupes de molécules. Et même un simple noyau atomique « bat » le temps puisqu’il existe une probabilité pour que le noyau fissionne de manière radioactive en produisant deux noyaux moins lourds et des radiations. Car un noyau atomique est déjà le siège de multiples rétroactions entre nucléons (protons et neutrons, quarks ainsi que leurs particules d’interaction, les gluons). Si la flèche du temps n’existe pas dans le vide quantique, elle se produit dès qu’il y a un nombre conséquent d’interactions de particules. Cela ne veut pas dire qu’il n’y ait aucun temps dans le vide mais un temps désordonné et sans flèche. Quant au Vivant, le développement précis de l’embryon avec la construction successive des différentes parties et organes du corps montre déjà l’existence d’une horloge produite par la rétroaction des gènes homéotiques. On a même montré qu’il existe des gènes qui, individuellement, sont des marqueurs de temps. Un simple groupe de cellules, comme les cellules de noyau suprachiasmatique de l’hypothalamus dans le cerveau qui comprend dix mille cellules pilotant vingt mille neurones, ou une protéine comme « Clock », pour le rythme circadien, peut produire, en rétroaction avec le matériel génétique comme facteur d’expression des gènes, un battement de temps. Comme le dit Rémy Lestienne dans « Les fils du temps », « La vie est un faisceau de cycles entrelacés »…
Boissin et Canguilhem dans « Les rythmes du vivant » :
« Un des grands problèmes commun aux animaux et aux végétaux est l’anticipation c’est-à-dire l’adaptation des êtres vivants à leur environnement par l’utilisation des horloges biologiques. »
Christian Vidal dans son article « Le chaos déterministe en chimie » :
« Lors de l’étude de cette réaction on s’est aperçu qu’une réaction chimique n’était susceptible d’osciller que si son mécanisme comporte une boucle de rétroaction. (...) L’effet renforce donc la cause qui lui donne naissance, c’est une auto-amplification Cette circonstance est rare, semble-t-il en chimie. Elle est beaucoup plus répandue dans le monde du vivant. (...) Les réactions biochimiques oscillantes sont peut-être la clé de la mystérieuse horloge interne qui réglerait les rythmes biologiques. On connaît depuis 1950, le caractère oscillant de certaines réactions du métabolisme. Parmi celles-ci la glycolyse est peut-être la plus importante (...) Elle constitue un mode essentiel et universel d’approvisionnement en énergie des cellules. Cette réaction oscille avec une période de l’ordre de la minute. (...) Il reste à comprendre comment les cellules d’un organisme parviendraient à coordonner leur activité pour former cette horloge biologique. »
Ilya Prigogine dans « La complexité, vertiges et promesses » :
« Le temps s’explique avec les bifurcations : le passé correspond à une trajectoire au travers de points de bifurcation et le futur comprend des bifurcations dont nous ne savons pas quelle sera la direction. »
Marc Bloch dans « Apologie pour l’histoire ou Métier d’historien » :
« Réalité concrète et vivante, rendue à l’irréversibilité de son élan, le temps de l’histoire, au contraire, est le plasma même où baignent les phénomènes et comme le lieu de leur intelligibilité. »
Jean-Pierre Changeux dans « L’homme neuronal » :
« Les organes des sens se comportent comme un « commutateur » d’horloges biomoléculaires. »
Stephen Jay Gould dans « Le pouce du panda » :
« L’histoire de la vie, telle que je la conçois, est une série d’états stables, marqués à de rares intervalles par des événements importants qui se produisent à grande vitesse et contribuent à mettre en place la prochaine ère de stabilité. Les organisme procaryotes (bactéries ou algues bleues) ont régné sur terre pendant 3 milliards d’années jusqu’à l’explosion cambrienne où la plupart des principales formes de vie pluricellulaires apparurent en l’espace de dix millions d’années. L’histoire de la terre peut être schématiquement perçue comme une série de pulsations occasionnelles forçant les systèmes récalcitrants à passer d’un état stable au suivant. »
Ilya Prigogine dans « Temps à devenir » :
« On a découvert que quand vous allez loin de l’équilibre, par exemple, en considérant une réaction chimique, que vous empêchez d’arriver à l’équilibre, se produisent des phénomènes extraordinaires que personne n’aurait cru possibles ; par exemple, des horloges chimiques. Une horloge chimique, qu’est-ce que c’est ? Prenons un exemple : vous avez des molécules qui de rouges peuvent devenir bleues. Comment imaginez-vous voir ce phénomène ? Si vous pensez que les molécules vont au hasard, vous allez voir des flashes de bleu, puis de flashes de rouge. Mais il se produit, loin de l’équilibre, dans d’importantes classes de réactions chimiques, des phénomènes rythmiques. Tout devient bleu, puis tout devient rouge, puis tout devient bleu, c’est-à-dire qu’une cohérence naît, qui n’existe que loin de l’équilibre. (…) Donc, loin de l’équilibre, se produisent des phénomènes ordonnés qui n’existent pas près de l’équilibre. Si vous chauffez un liquide par en-dessous, il se produit des tourbillons dans lesquels des milliards de milliards de molécules se suivent l’une l’autre. De même, un être vivant, vous le savez bien, est un ensemble de rythmes, tels le rythme cardiaque, le rythme hormonal, le rythme des ondes cérébrales, de division cellulaire, etc. Tous ces rythmes ne sont possibles que parce que l’être vivant est loin de l’équilibre. Le non-équilibre, ce n’est pas du tout les tasses qui se cassent ; le non-équilibre, c’est la voie la plus extraordinaire que la nature ait inventée pour coordonner les phénomènes, pour rendre possibles des phénomènes complexes. »
Ladislas Robert dans « Le temps et sa flèche » :
« Pratiquement toutes les manifestations biologiques suivent des rythmes. Certains rapides, d’autres lents. mais pratiquement aucun phénomène biologique ne se déroule selon une cinétique continue et linéaire… Les enzymes consommant de l’énergie (ATP) seraient activées à des valeurs élevées en charge d’adeylique, par l’ATP et inhibées par un excès d’ADP et AMP. (...) Ces oscillations métaboliques pourraient représenter une des horloges élémentaires de la cellule. Il apparaît que ces oscillations sont à leur tour "entraînées" par d’autres rythmes, plus lents caractérisant le domaine épigénétique de la cellule - biosynthèse des macro-molécules, leurs interactions - comme le suggère les travaux sur les amibes de Lloyd et Edwards. (...) Le couplage étroit des réactions rapides et lentes, médié par des cofacteurs et métabolites intermédiaires permet ainsi à la cellule d’ajuster ses oscillations métaboliques à la demande énergétique au lieu de suivre sa capacité maximale de génération d’énergie. (...) Certaines horloges peuvent être synchronisées, "entraînées" sur un rythme compatible avec la survie optimale de la cellule ou de l’organisme, d’autres phénomènes suivent un rythme fortement dépendant de facteurs intrinsèques comme par exemple l’utilisation du glucose pour générer l’énergie indispensable à la vie ainsi que son action délétère sur les protéines de structure. »
Jean Chaline dans « Les horloges du Vivant » :
« Le nouveau stade de la théorie de l’évolution prend donc en compte le fait que le vivant porte en lui les potentialités de son évolution grâce au jeu de plusieurs mécaniques, celles des mutations et du développement qu’elles conditionnent ; un véritable bricolage biologique contrôlé par la sélection naturelle. Mais ces contraintes internes interfèrent avec les contraintes externes de l’histoire de la Terre et des modifications de l’environnement qui introduisent la contingence dans l’évolution ; c’est le poids de l’histoire qui imprime fortement sa marque à l’évolution. Les qualificatifs hiérarchique, saltatoire, événementiel, contingent et chaotique expriment la nouvelle dimension globale de la théorie de l’évolution et la rapprochent des théories des sciences physiques expliquant la constitution et l’évolution générale de l’univers… On peut dire que les hétérochronies du développement, qui expriment la dynamique de l’évolution spatio-temporelle des structures, opèrent à la façon de véritables horloges internes contrôlant le développement de chaque caractère. Les mutations dérèglent le fonctionnement des horloges ancestrales en le ralentissant, l’accélérant, le retardant ou l’avançant. (…) Cette nouvelle conception s’oppose complètement à celle du stade synthétique de la théorie où l’on donnait à l’environnement et à la sélection naturelle le rôle primordial. (…)Les résultats actuels des recherches entreprises sur le rôle des gènes architectes moléculaires universels de la famille des gènes à homéobox ou « homéogènes », bien que partiels et préliminaires, constituent une véritable révolution de fond dans la compréhension du phénomène évolutif… Précisons tout d’abord la signification du mot « homéogène » qui traduit le fait que leur mutation transforme un segment du corps d’un insecte en un autre segment ; « homéo » signifiant semblable. Ces gènes, désignés tout d’abord sous le nom de gènes HOM chez les invertébrés et Hox chez les vertébrés, sont maintenant réunis sous le seul vocable de gènes Hox. On s’est en effet aperçu que ces gènes étaient très semblables pour la bonne raison que les gènes Hox des vertébrés proviennent de duplications des gènes HOM des invertébrés. En effet, le remplacement de gènes HOM de la drosophile par des gènes Hox de souris ne perturbe pas leur développement… On sait maintenant que les homéodomaines de la drosophile et de la souris sont presque identiques. Par exemple, l’homéodomaine Antennapedia de la drosophile ne diffère de celui du gène HoxB6 de la souris que par quatre acides aminés sur soixante et un… Les homéogènes constituent les ressorts des horloges internes du vivant. »
Ladislas Robert dans « Les temps de la vie » :
« Une famille de gènes, les homéogènes, comportent des messages plus compliqués concernant le plan d’organisation de l’animal. Apparus chez les invertébrés, bien étudiés chez les insectes (la mouche de vinaigre ou drosophile) et les vers plats (caenorhabditis elegans), on les retrouve chez les vertébrés jusqu’à l’homme et l’on perce progressivement le secret de leur fonctionnement. Un aspect majeur de leur action reste cependant obscur : leur capacité à assurer un chronométrage précis de la mise en place des parties du corps… Les recherches très actives devraient nous permettre d’approfondir l’horlogerie des gènes du développement dans un avenir proche… L’équipe de Mme Le Douarin a identifié un gène, appelé « c-hairy 1 » qui correspond au gène de même fonction appelé « hairy » chez la mouche drosophile et dont l’expression rythmée dans le temps semble régler la vitesse d’apparition des somites (segments du corps de l’embryon). Ce gène code un facteur de transcription (des protéines agissant sur des gènes qui activent ou au contraire répriment leur expression)… L’identification de cette horloge cellulaire concernant une partie importante de l’embryon incite à penser que la découverte d’autres horloges cellulaires dictant le rythme du développement des autres parties de l’embryon est possible. La connaissance de ces horloges cellulaires est complétée par celle des gènes réglant la construction spatiale de l’embryon. Le fonctionnement de ces homéogènes, identifiés il y a à peine quelques décennies, a révélé le plan de construction de l’embryon, l’espacement des segments et des organes de l’embryon, selon les axes symétriques antéro-postérieur et dorso-ventral. (…)
Britton Chance, biologiste américain travaillant à Philadelphie, a pu réaliser des observations importantes confirmant la rythmicité des phénomènes métaboliques, c’est-à-dire leurs variations d’intensité dans le temps… En utilisant une suspension de cellules de levure, il montre que les oscillations de la glycose peuvent se maintenir pendant des périodes longues. Observation surprenante, ces oscillations persistent quand on enregistre le phénomène non plus avec des cellules intactes, mais en utilisant un extrait de cellules, donc en l’absence de cellules vivantes. La nature discontinue, oscillante, de la glycose n’est donc pas la propriété de la seule cellule vivante, mais est conservée dans un extrait qui contient l’ensemble des biocatalyseurs qui en sont responsables… Pour observer les oscillations dans un extrait cellulaire, il fallait attendre la découverte du rôle clef de certaines enzymes dans ces chaînes de réactions qui, grâce à leurs propriétés « allostériques » (modifications de conformations qui modifie leurs activités catalytiques) régulent le débit de la chaîne de réactions. Concept développé par Jacob, Monod et Changeux à Paris et par Arthur Pardee aux Etats-Unis, il s’agit de la régulation d’une chaîne de production biochimique par la rétroaction d’un des produits de cette réaction sur un catalyseur situé au début de la chaîne. Ce concept s’est avéré critique pour la compréhension des réactions métaboliques ; il permet d’expliquer certains de ces phénomènes oscillatoires observés au cours du métabolisme cellulaire. En l’occurrence, c’est l’enzyme appelée phosphofructokinase (PFK) qui possède cette propriété allostérique… En court-circuitant PFK, on supprime les oscillations… (...) Le plus souvent, les produits d’une réaction inhibent l’enzyme allostérique qui régule le débit de la chaîne métabolique (...) Dès que la quantité du produit final baisse, la chaîne de production peut redémarrer (...). Ce type de coopérativité entre site catalytique et site régulateur joue un rôle important dans la coordination de nombreuses réactions métaboliques dans la cellule. (...) Le modèle permettant de décrire quantitativement le comportement de cette enzyme a été proposé par Goldbeter… Les calculs théoriques de Goldbeter permettent de représenter les oscillations entretenues du système… En variant la concentration du substrat, le système évolue vers un cycle limite (conformément aux théorèmes de Poincaré, Bendixson, Nicolis et Prigogine). La représentation de cette évolution vers un cycle limite dans l’espace de phase correspond à un « attracteur étrange ». Ce formalisme, dit du « chaos déterministe », est en effet applicable à de nombreuses réactions biologiques… Ce même type de comportement oscillant a aussi été observé dans les cellules musculaires. La périodes oscillations varie avec la température, elle augmente si la température baisse. Britton Chance suggère que ces oscillations pourraient être à l’origine des rythmes circadiens… Le cas évoqué plus haut de la glycose nécessite des intervalles de temps qui se comptent en secondes ou en minutes… D’autres réactions beaucoup plus rapides comme la fixation d’un ligand sur une protéine ou les modifications de l’acidité du milieu se jouent en millisecondes. C’est le cas aussi des réactions d’oxydoréduction qui se déroulent dans les mitochondries au cours du métabolisme oxydatif…
Le réglage des horloges circadiennes
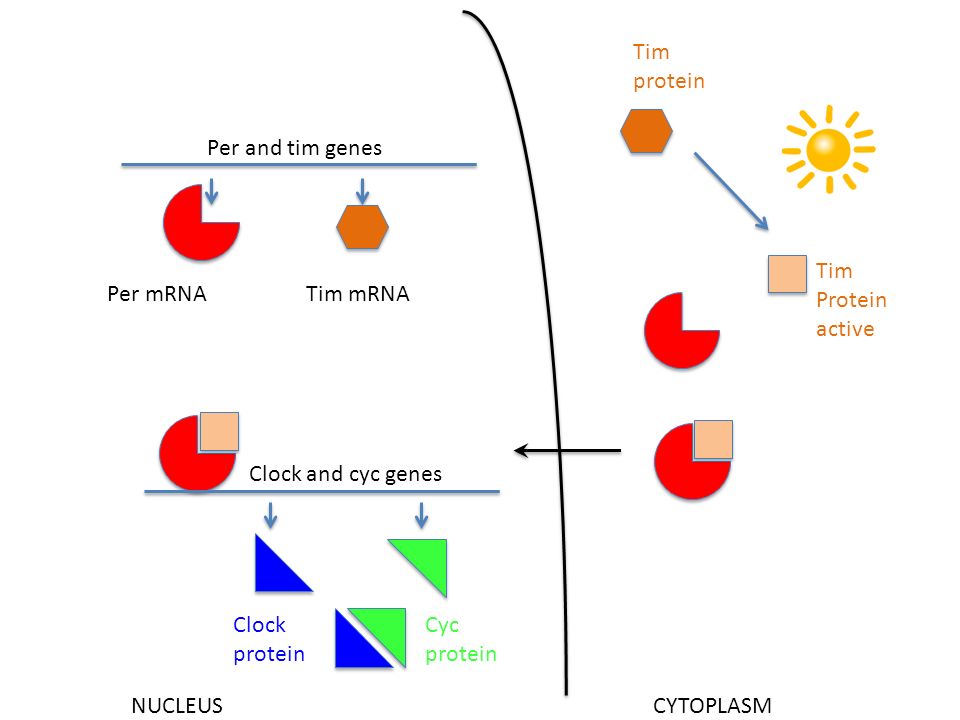
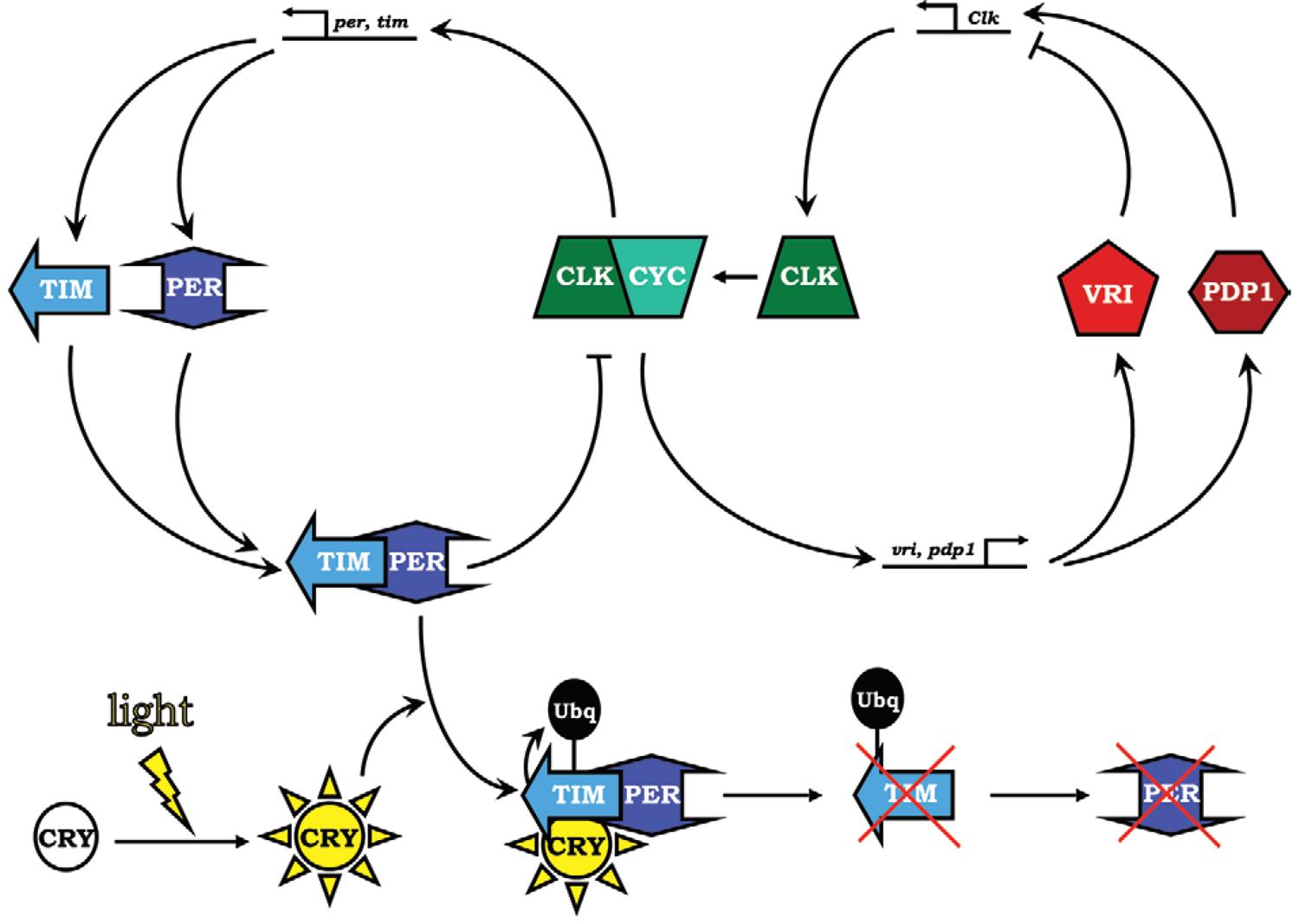
Les observations précoces des botanistes ont vite été suivies par celles du règne animal, puis par celles de l’homme. Karl von Frisch fut l’un des premiers à observer le fonctionnement d’une horloge chez les invertébrés… Entre l’observation qui permet de conclure à l’existence d’horloges internes et la reconnaissance explicite du fait, il s’est écoulé presque un demi-siècle. On attribue à Gustav Kramer et Klaus Hoffmann la démonstration des horloges internes… Au cours de la deuxième moitié du XXe siècle, plusieurs équipes ont cherché à identifier le codage génétique des horloges biologiques… Chez l’homme et, peut-être, chez des vertébrés, le centre nerveux appelé « noyau supraschiasmatique », localisé juste au-dessus du croisement des nerfs optiques ou rétiniens, semble jouer un rôle important sans pour autant représenter le seul et unique contre chronologique de l’organisme… Si l’on transplante le noyau supraschiasmatique d’animaux jeunes à des animaux âgés… on « rajeunit » le comportement rythmique de l’animal…. Michael W. Young, à l’Institut Rockfeller de New York, a pu reconstituer avec des mouches à vinaigre le phénomène du décalage horaire… L’arrivée de la lumière sur la rétine de ces mouches provoque la dégradation d’un complexe macromoléculaire composé de deux protéines, appelées PER et TIM, codées par deux gènes (per et tim). Ces protéines font partie d’un circuit de rétrocontrôle inactivant les deux gènes qui codent leur séquence. Vers le milieu de la journée, le complexe protéique PER-TIM est dégradé et c’est un autre complexe, composé de deux protéines, CYCLE et CLOCK, qui se fixe sur les gènes per et tim pour les inciter à se réexprimer et produire l’ARN messager permettant la resynthèse des protéines PER et TIM. Leur complexe se reconstitue au coucher du soleil et commence à freiner le fonctionnement des gènes cycle et clock qui code les deux protéines éponymes, dont le complexe active l’expression des gènes per et tim. Le complexe protéique PER-TIM supprime ainsi, dès le coucher du soleil et pendant la nuit, sa propre production par la répression des gènes qui codent ces protéines… Ce mécanisme de tic-tac moléculaire a pu être confirmé, avec quelques variantes et selon une complexité accrue, de la mouche à l’homme… On constate alors l’expression oscillante de ce gène-horloge, conformément au schéma proposé pour la drosophile… Ces gènes de l’horlogerie – per, tim, cycle et clock, notamment -, impliqués dans le réglage de l’horloge « centrale », sont présents dans toutes les cellules de l’organisme, en plus des neurones et de cellules gliales du noyau supraschiasmatique. Ces gènes sont-ils réprimés dans les très nombreuses autres cellules de l’organisme ou bien peuvent-ils s’exprimer et jouer un rôle d’horloge secondaire ? (…) L’équipe américaine de Stevens Kay à la Scripps Clinique en Californie a mis en évidence ce type d’horlogerie au niveau d’autres organes de la mouche, dans les ailes, les pattes, autour de la bouche ainsi que dans les antennes… Ce type de fonctionnement extracérébral des gènes mentionnés se retrouve chez les vertébrés : dans les fibroblastes, les cellules ubiquitaires des tissus conjonctifs, le système d’horloge fonctionne aussi d’une façon cyclique. »
Arthur Winfree dans « Les horloges de la vie » :
« Rien ne nous semble plus ordinaire que l’alternance du sommeil et de la veille, qui rythme inlassablement notre vie. Tandis que tourne la Terre, notre conscience poursuit ses propres révolutions, passant du rêve, mystérieux et solitaire, à l’éveil, avec ses activités sociales et professionnelles. A l’occasion, nous avons tous modifié le rythme quotidien de nos activités et nous savons que notre rythme interne n’est pas strictement synchronisé sur l’alternance du jour et de la nuit. Nous savons également que nous ne pouvons échapper au sommeil bien longtemps. Le rythme de l’horloge géophysique est inscrit en nous ; ce n’est pas seulement le fruit d’une longue habitude, contractée dès la naissance, mais un héritage physiologique. (…) La température interne varie imperturbablement, avec une période de 25 heures (appelé rythme circadien par opposition au rythme diurne de 24 heures – circadien veut dire proche du rythme des jours). En découvrant la régularité du cycle des températures internes, Jurgen Aschoff, directeur de l’Institut Max Planck d’Andechs, a ainsi démontré l’existence d’une horloge circadienne interne. La période de l’alternance sommeil-veille d’un homme est de 24 heures mais elle se poursuit selon sa période naturelle, égale à 25 heures, quand l’homme a été isolé, soustrait à l’influence de la lumière. (….) On ne connaît le rythme interne de la période que depuis 20 ans. D’autres primates possèdent également ce rythme. (…)Tout notre environnement varie selon un rythme « civil » de 24 heures et nos fonctions physiologiques accélèrent ou ralentissent ainsi de concert avec le cycle du sommeil. Les conséquences médicales de cet asservissement sont aujourd’hui connues. (…) On supporte mieux la douleur l’après-midi que le matin. Les anesthésies sont particulièrement efficaces en début d’après-midi et les doses nécessaires le matin peuvent être excessives en fin de journée. Les sujets allergiques sont beaucoup plus sensibles juste après minuit que dans l’après-midi. Le foie métabolise bien plus vite l’alcool en début de soirée que le matin. Les médecins doivent tenir compte des rythmes biologiques pour l’interprétation des examens biologiques et l’établissement des diagnostics. Une température égale à 37 degrés pendant la nuit est anormale ;l à trois heures du matin, cette valeur indique que le patient est fiévreux, alors qu’en fin d’après-midi, elle n’est pas inquiétante. (…) En fait, c’est non seulement pour le diagnostic, mais aussi pour le traitement que le médecin doit tenir compte des rythmes circadiens. Comme la réplication de l’ADN, dans de nombreux types de cellules, s’effectue de préférence à certaines heures de la journée, les radiothérapies ou les chimiothérapies sont beaucoup plus efficaces à des moments précis de la journée. On observe parfois que 80% des cellules d’une population résistent au traitement à un moment de la journée alors que la même dose est fatale à 80% de la population quelques heures plus tard. (…) Les endocrinologues et les vétérinaires connaissent depuis longtemps l’importance de l’heure d’administration des hormones. On traite les insuffisances surrénaliennes humaines, par exemple, en administrant plutôt la cortisone le matin lorsque l’activité de la glande cortico-surrénale est maximale. (…) On a également observé que les troubles du sommeil et diverses maladies psychiatriques plus graves, notamment des dépressions, sont synchronisées avec la libération d’hormones dans le cerveau : ces maladies, jusqu’ici incurables, pourraient être des effets secondaires de troubles des rythmes circadiens ; des expositions convenablement programmées à la lumière (celle du soleil ou celle de lampes intenses) permettent de traiter certaines formes de dépression). On suppose que l’exposition à la lumière modifie la sécrétion de mélatonine, une neuro-hormone associée au cycle circadien. (…) Bien que le rythme naturel de notre horloge soit plus lent d’une heure par jour environ que le rythme de rotation de la Terre, notre horloge est entraînée par les variations lumineuses et sa période est ainsi réglée sur 24 heures. Un enfant possède d’abord un rythme naturel de période égale à 25 heures, puis il s’adapte au rythme de son entourage. (…) Une horloge ne sert à rien si on ne peut la remettre à l’heure ; de même une horloge biologique qui ne se resynchroniserait pas serait inutile, sauf si sa période était exactement celle de la Terre. Elle devrait en outre être insensible au froid, au chaud, aux émotions et aux variations hormonales. La moindre différence entre sa période interne et la période externe la mettrait hors d’usage : même si le décalage était seulement d’une minute par jour les animaux diurnes que nous sommes seraient, en deux ans, des animaux nocturnes s’éveillant au crépuscule, se couchant à l’aube : deux ans plus tard, nous serions à nouveau en phase avec le Soleil et, deux ans plus tard encore, nous serions à nouveau plongés dans la nuit. (….) Pour demeurer accordés à l’environnement, il nous faudrait vivre en nomades, et faire le tour de la Terre. (…)Pour rester synchronisée, notre horloge interne doit battre à un rythme peu différent de celui de la Terre et se recaler quotidiennement. Cette possibilité de resynchronisation est essentielle au fonctionnement de toute horloge biologique : l’animal qui la possède demeure ainsi dans sa zone horaire d’origine sans devoir voyager pour compenser l’inévitable décalage. La resynchronisation résulte d’un entraînement. Un cycle est ainsi entraîné par un autre cycle, grâce à des signaux réguliers, à la façon des danseurs qui suivent le rythme d’une musique. Le cycle des saisons, par exemple, entraîne celui de la floraison et de la fructification de nombreux arbres ; la pousse des bois du cerf est soumise à la même régulation. Dans le cas des rythmes circadiens, le signal de commande est généralement la durée quotidienne du jour. Selon les cas, la lumière retarde ou avance légèrement l’horloge interne. Il existe bien d’autres systèmes de resynchronisation des horloges biologiques, mais la grande majorité des espèces vivantes se fondent sur l’événement quotidien qui se répète le plus régulièrement : l’alternance du jour et de la nuit ; c’est à ce signal que leurs horloges circadiennes sont devenues les plus sensibles. (…) Comment s’effectue cette resynchronisation ? (…) Les stimuli (une exposition à la lumière blanche pendant 14 heures, par exemple) agissent différemment selon la phase du cycle interne à partir duquel on les applique. En première approximation, ces stimuli n’engendrent qu’un décalage : une période après le début de la stimulation, l’horloge n’est plus à la phase qu’elle aurait eu sans la stimulation, mais à une nouvelle phase. (...) On rencontre souvent en physiologie le déclenchement d’un événement discontinu, par exemple le passage du sommeil à l’état de veille, qui résulte d’un processus comportant un phénomène de seuil et que ce seuil soit lui-même le produit d’un rythme. Le fonctionnement passe par une phase critique. En termes de rythmes, le passage d’une resynchronisation paire à impaire et inversement est une discontinuité fondamentale. (…) Tous les organismes subissent une resynchronisation impaire lorsque les stimuli sont très faibles ; à la limite (stimuli nul), la nouvelle phase est égale à l’ancienne. Ces mêmes organismes ont-ils une resynchronisation impaire quand on les stimule davantage ? (…) Quand la durée d’exposition périodique à la lumière passe de 7,5 minutes à 2 heures, (…) la resynchronisation d’impaire devient paire. Ce changement n’a pas seulement lieu quand on multiplie par 16 la durée de stimulation : on pourrait le cerner bien plus précisément en augmentant progressivement la durée des stimulations dans diverses expériences. Cette simple remarque suffit à réfuter l’idée selon laquelle les différents types topologiques de resynchronisation résulteraient de mécanismes d’horloge différents ou de types de stimuli particuliers. Au contraire, les différentes resynchronisations semblent dues à un mécanisme universel ! Cette observation fait surgir un nouveau problème : quel que soit l’organisme considéré et quelle que soit la phase à laquelle on le stimule, (…) le passage d’une resynchronisation impaire à une resynchronisation paire, c’est-à-dire le passage d’une boucle autour du trou central du tore (représentation de l’espace des états sur un équivalent topologique de la chambre à air) à une boucle ne traversant pas ce trou, ne peut pas s’effectuer de manière continue. (...) Pour les moustiques Culex, par exemple, le renforcement de la stimulation finit par modifier brusquement la courbe de resynchronisation : tandis qu’elle décrivait toutes les valeurs de la nouvelle phase, un intervalle complet de valeurs est maintenant épargné. A partir d’une valeur critique de la phase, lorsque le stimulus est assez intense, la nouvelle phase décroche. Le mot "décroche" peut sembler abusif car il faut imaginer une discontinuité et, hormis les événements moléculaires ou relevant de la mécanique quantique, les changements temporels (en biologie) semblent continus, bien que parfois très rapides. Les véritables discontinuités (en biologie) sont des phénomènes rares et inobservables. Comment prouver que la cause est réellement inférieure à n’importe quelle valeur finie (c’est-à-dire prouver la continuité) ? Pourtant dans le cas présent, la nouvelle phase semble bien changer d’une quantité finie, alors que l’ancienne phase ne change que d’une façon infinitésimale. Serions-nous dans le cas exceptionnel d’une véritable discontinuité ? Oui, car, dans le phénomène considéré, la discontinuité est un impératif topologique. Pour transformer une boucle entourant le trou d’un tore en une boucle ne l’entourant pas, il faut nécessairement déformer la boucle de façon discontinue. Lorsque le stimulus atteint la valeur critique, la boucle doit être coupée, déplacée, puis recollée. La topologie ne décrit pas la discontinuité, mais elle prévoit un phénomène à découvrir : il doit exister une singularité physiologique. Il y a 20 ans seulement, on ne soupçonnait même pas l’existence de cette singularité, mais les expériences où l’on observait un déphasage variant continûment en fonction des conditions expérimentales auraient dû nous mettre sur la voie de la singularité inobservée. La topologie a indiqué la méthode à suivre pour trouver expérimentalement le point singulier et, rapidement, le protocole de piégeage de singularité a été mis en œuvre pour la recherche de la phase singulière d’une horloge biologique. (…) L’examen raisonné des resynchronisations observées suffit à déduire les propriétés essentielles des horloges biologiques. Cet examen est favorisé par une présentation géométrique, intuitive, fondée sur des diagrammes de phase colorés. Sur ces diagrammes apparaissent directement les surprenantes conséquences de la découverte des resynchronisations paires. (…) A la limite de la resynchronisation impaire, la nouvelle phase est strictement égale à l’ancienne ; à la limite de la resynchronisation impaire, la nouvelle phase est constante, quelle que soit l’ancienne phase. (…) Construisons alors un graphique rectangulaire où apparaissent tous les types de stimulations possibles : en abscisses, nous portons les anciennes phases, à partir desquelles les horloges sont stimulées, en ordonnée, nous portons les intensités de stimulation. En chaque point de ce rectangle, nous indiquons le résultat de l’expérience (la nouvelle phase) par un code de couleur. (…) Représentons la phase nulle par la couleur rouge et convenons que les phases croissantes soient successivement représentées par le violet, le pourpre, le bleu, le vert, le jaune, l’orange et de nouveau le rouge. (…) Selon l’hypothèse de continuité, la couleur en chaque point doit être voisine de la couleur des points voisins. Une zone jaune, par exemple, ne peut être voisine d’une zone bleue : entre les deux doit apparaître soit une zone verte, soit une succession de zones orange, rouge, pourpre, violette. (…) Faites l’expérience, vous n’éviterez pas une zone où toutes les couleurs se mélangent. Cette conclusion est biologiquement importante : pour un point correspondant à un couple précis d’ancienne phase et d’intensité du stimulus, la nouvelle phase est indéterminée, bien que les points très voisins soient de toutes les couleurs. Ce point est une singularité de phase. »
Références :
1. Arthur Winfree Les horloges de la vie : les mathématiques des rythmes biologiques Pour la science (BELIN) 1997
2. Arthur Winfree The geometry of biological time. Springer-Verlag 1980
3. Steven Strogatz SYNC : the emergeous science of spontaneous order. Hyperion 2003
4. Michèle Teboul et Franck Delaunay Ni maître ni esclave chez les horloges biologiques Médecine-Sciences Juillet-Aout 2004
5. Hugues Dardente. Redondance génétique et synchronisation cellulaire dans les horloges circadiennes. Medecine-Sciences. Mars 2008.
6- Albert Goldbeter. La vie oscillatoire.
7- Gilbert Simondon – Les rythmes comme cycles de l’ontogenèse ?
Albert Goldbeter écrit dans « La vie oscillatoire » :
« Battements du cœur, respiration, alternance des phases d’éveil et du sommeil, cycle ovarien, migrations animales, floraison des plantes : la vie est rythmes. (…) De nombreux processus périodiques sont à l’œuvre, de manière imperceptible au sein de nos cellules. Ainsi, une horloge biologique contrôle le cycle de division cellulaire, des premiers stades de l’embryon jusqu’à la formation de l’organisme adulte au sein duquel elle poursuit son action tout au long de la vie. L’action des neurones est sous-tendue par des rythmes. D’autres exemples nous sont donnés par l’alternance périodique des différentes phases du sommeil, les nombreux rythmes hormonaux et les oscillations dans les communications intercellulaires. (…) Parmi les multiples rythmes qui scandent la physiologie des organismes, certains sont liés à la périodicité de l’environnement. Ainsi des rythmes circadiens, d’une période proche de 24 heures, permettent aux êtres vivants de s’adapter à l’alternance du jour et de la nuit, résultent de la rotation de la Terre sur son axe. D’autres rythmes sont liés au cycle annuel des saisons, comme la floraison, l’hibernation, les migrations ou la reproduction chez de nombreuses espèces animales. (…) Les rythmes biologiques les plus rapides caractérisent l’activité électrique des cellules nerveuses ou musculaires. Leur période peut varier du centième de seconde à une dizaine de seconde ou plus. (…) Le cerveau est un organe rythmique par excellence, tout comme le cœur. (…) Les mécanismes qui sous-tendent la périodicité cyclique au niveau cellulaire sont similaires aux mécanismes des rythmes neuronaux. A côté des oscillations d’origine électrique qui forment une classe majeure parmi les rythmes biologiques, il en existe de nombreux autres de nature non électrique. Ainsi, des oscillations sont observées dans les voies de communication entre cellules, par exemple de nature hormonale, et dans le cycle de division cellulaire dont la période varie d’une dizaine de minutes pour certaines cellules embryonnaires à 24 heures ou plus pour les cellules de l’organisme adulte. D’autres rythmes sont observés au sein de la cellule. Ainsi, des oscillations de calcium surviennent avec une période de quelques secondes ou de quelques minutes dans des cellules stimulées par une hormone ou un neurotransmetteur. (…) Un bel exemple de rythme dans les communications intercellulaires est fourni par des amibes dites « sociales », qui communiquent entre elles par des signaux pulsatiles dont la fréquence est de l’ordre d’une pulsation toutes les 5 ou 10 minutes. De même, la plupart des hormones sont sécrétées de manière pulsatile avec une périodicité allant d’une dizaine de minutes pour l’insuline à une heure pour l’hormone qui induit la sécrétion des hormones gonadotropes impliquées dans le contrôle de la reproduction et de trois à cinq heures pour l’hormone de la croissance. (…) D’autres rythmes procèdent d’un mécanisme supracellulaire faisant intervenir des interactions régulatrices entre les différents organes. Ainsi, cycle ovarien, chez la femme, prend la forme du cycle menstruel d’une période proche de 28 jours et fait intervenir les sécrétions hormonales de l’hypothalamus, de l’hypophyse et des ovaires. A un niveau écologique dépassant celui des organismes, les interactions entre populations animales donnent lieu à des oscillations de période encore plus longue. (…) Les oscillations dans les populations de prédateurs et de proies représentent un exemple classique de rythme en écologie, avec une période pouvant atteindre plusieurs années. (…) Parmi les rythmes multiannuels remarquables, ceux dont la période est la plus longue dans le règne animal caractérisent le cycle de la vie de certaines cigales qui émergent du sol, dans l’Est des Etats-Unis, tous les 13 ou 17 ans. Enfin, dans le règne végétal, certaines espèces de bambou fleurissent avec une périodicité qui peut atteindre la centaine d’années. Dans la plupart des exemples connus expérimentalement la non-linéarité responsable de l’existence d’oscillations est associée à la présence d’une étape de nature auto-catalytique, c’est-à-dire auto-amplifiée, en raison d’une rétroaction positive. Souvent des rétroactions positives et négatives entremêlées sont à la base du comportement oscillant. Ces conditions d’ouverture et de non-linéarité sont précisément celles qui caractérisent la chimie des êtres vivants. (…) Une observation parmi les plus énigmatiques au sujet des rythmes circadiens est la possibilité de les supprimer de manière permanente par une brève impulsion lumineuse. Tout aussi mystérieuse est la restauration du rythme par une seconde impulsion identique à la première. Ce phénomène, observé chez la drosophile et chez certains mammifères comme le cochon d’Inde, (…) a reçu une interprétation par Arthur Winfree. (…) Winfree suppose que les rythmes circadiens se produisent sous forme d’oscillations de type cycle limite autour d’un état stationnaire instable. L’effet de la perturbation lumineuse est de ramener chaque cellule oscillante à son état stationnaire. Il est nécessaire pour cela que la perturbation possède une amplitude précise et soit appliquée à une phase particulière des oscillations. Comme l’état stationnaire est instable, les cellules finissent par le quitter et retournent chacune au cycle limite. Toutefois, la phase des oscillations est indéfinie, et chaque cellule rejoint le cycle à une phase aléatoire. La désynchronisation qui en résulte abolit l’émergence d’un rythme circadien global, au moins pendant un certain temps. L’effet de la perturbation lumineuse pendant ce laps de temps est de supprimer le rythme circadien. (…) Le but premier de ce livre a été de rechercher le mécanisme à l’origine des principaux rythmes du vivant. La plupart du temps, ce mécanisme est de nature cellulaire et repose sur des processus de contrôle impliquant la régulation de gènes, de récepteurs, de canaux ioniques ou d’enzymes. (…) Pour que le rythme s’établisse de manière robuste au niveau d’un tissu, d’un organisme ou d’une population, il faut encore que les oscillateurs se synchronisent entre eux. (…) Nous avons rencontré de multiples exemples de synchronisation des rythmes : oscillations glycolytique dans une population de levures, agrégation périodique des amibes sociales, mise en phase d’oscillateurs neuronaux sous-tendant l’activité rythmique du cerveau, sécrétion coordonnée des pulsations d’insuline par les cellules béta du pancréas, émergence du rythme cardiaque au sein des cellules du nœud sinusal et des rythmes circadiens dans des cultures des cellules des noyaux supraschiasmatiques. Dans chacun de ces exemples, des cellules oscillantes se synchronisent entre elles. (…) Le couplage et la synchronisation jouent un rôle clé dans la dynamique des systèmes du vivant. »
Jean-Didier Vincent, dans la préface de « Les fils du temps » de Rémy Lestienne :
« Il existe une horloge formée d’une centaine de neurones dans l’œil de l’escargot : elle indique les jours avec une précision digne d’un horloger helvète. On dit qu’il s’agit d’un « pacemaker » circadien parce qu’elle a la propriété de produire des rythmes à la façon d’un métronome battant la mesure selon une période d’environ un jour. Ce rythme est une propriété intrinsèque des cellules oculaires ; isolées de l’œil et du reste du corps, celles-ci continuent de présenter une alternance d’activité et de repos selon une périodicité de vingt-quatre heures. Comme toute horloge, la montre vivante doit être remise à l’heure en fonction des saisons et des fuseaux horaires. C’est le cerveau qui réalise ce réglage par l’intermédiaire des voies nerveuses qui changent la phase de l’horloge sans en modifier la période. Pour transmettre ses ordres, il utilise un messager chimique, la sérotonine, véritable horloger du vivant présent chez toutes les espèces d’animaux supérieurs. Fabriquer du temps semble donc être une propriété fondamentale du vivant. Le rythme circadien constitue une formidable possibilité d’adaptation aux conditions de vie à la surface de la Terre. Tous les êtres vivants plus évolués que les bactéries présentent des rythmes biologiques (alternance d’activité-repos, cycle des températures, etc.) qui se produisent avec une périodicité de vingt-quatre heures. Ces rythmes circadiens sont endogènes et font partie intégrante de la cellule. Chez les êtres pluricellulaires, seules quelques cellules conservent cette propriété de produire un rythme circadien. Ce sont les horloges internes, constituées de neurones spécialisés, que possèdent non seulement l’escargot mais aussi des animaux plus évolués : le rat, le lapin… et l’homme. Personne ne peut échapper à ces cadences imposées par le cerveau… Nombre de nos activités, de nos sécrétions hormonales, de mouvements de nos viscères ne pourraient pas se passer de ce temps quotidien. De multiples « montres » logées dans notre cerveau règlent l’emploi du temps de chacune de nos fonctions. Il semble qu’une seule horloge, dite « maîtresse », coiffe et gouverne cet amoncellement de pendules, un peu comme à la gare Saint-Lazare. Elle est enfouie au plus profond du cerveau, dans le « noyau suprachiasmatique » de l’hypothalamus. Le rythme circadien existe déjà chez le fœtus : il est alors entravé par celui de la mère, dont il peut toutefois être facilement dissocié. Les plans de l’horloge font partie du patrimoine génétique de l’individu. Des biologistes ont récemment isolé, sur le chromosome X de la mouche drosophile, le gène responsable de cette périodicité. »
Lire aussi :
Gènes architectes et horloges du développement
Les horloges du Vivant, Jean Chaline
Les horloges du vivant, André Klarsfeld
Les rythmes biologiques, un processus dynamique et des structures auto-organisées et émergentes
Albert Goldbeter, La vie oscilatoire. Au cœur des rythmes du Vivant
Ouvrage collectif, Le temps du Vivant
Jean-Nicolas Tournier dans Le temps et sa flèche
Exposition « Les horloges du Vivant »
Cité des Sciences, Horloge biologique : au rythme de l’ADN
Clock : les Horloges du Vivant
Les rouages de l’horloge biologique
Les mécanismes moléculaires de l’horloge circadienne
Comment la protéine Clock rythme notre biologie
Conférence :