Accueil > 02 - Livre Deux : SCIENCES > Développement et génétique > La vision produite par œil-nerf optique-cerveau, un produit de la (...)
La vision produite par œil-nerf optique-cerveau, un produit de la génétique, du développement et de l’évolution, vue… par des grands scientifiques, de Darwin à Gould en passant par Gehring et Feynman
lundi 14 mai 2018, par
« Pouvons-nous croire que la sélection naturelle puisse produire, d’une part, des organes insignifiants tels que la queue de la girafe, qui sert de chasse-mouches et, d’autre part, un organe aussi important que l’œil ? (…)Il semble absurde au possible, je le reconnais, de supposer que la sélection naturelle ait pu former l’œil avec toutes les inimitables dispositions qui permettent d’ajuster le foyer à diverses distances, d’admettre une quantité variable de lumière et de corriger les aberrations sphériques et chromatiques. (…) La difficulté d’admettre qu’un œil complexe et parfait a pu être produit par la sélection naturelle, bien qu’insurmontable pour notre imagination, n’attaque en rien notre théorie. »
Darwin, « L’Origine des espèces »
L’œil a été longtemps présenté par les créationnistes comme la preuve de la fausseté de la théorie de l’évolution de Darwin et comme l’incontournable nécessité du créationnisme. Le fait que quarante fois quelque chose d’aussi parfait que l’œil ait pu être fabriqué pourrait sembler la meilleure preuve de la création divine. Et pourtant, les dernières découvertes des sciences ont tranché cette controverse. Lire ici l’argumentaire sur l’œil des créationnistes contre Darwin

On a d’abord cru que des yeux différents de diverses espèces avaient des origines différentes mais...
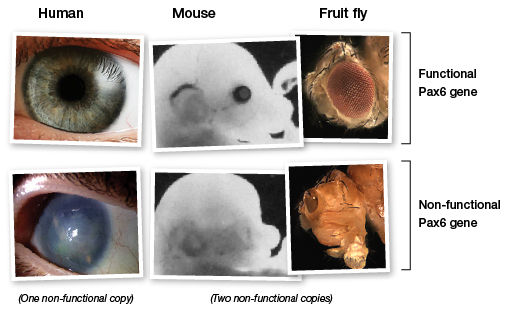
Malgré l’existence d’au moins quarante type d’yeux chez les animaux, les gènes homéotiques nous donnent aujourd’hui la preuve que tous ces yeux, aussi divers soient-ils de forme, de mode d’organisation, de fonctionnement, de plan, ont une seule et même origine et ont évolué à partir de celle-ci comme le pensait Darwin…
Pour toutes les espèces ayant un oeil, du poisson à l’insecte en passant par le mammifère, celui qui a produit l’oeil (le gène régulateur maître de la production de l’oeil dans le développement de l’animal) s’appelle le gène Pax-6. Et voici sa protéine :






















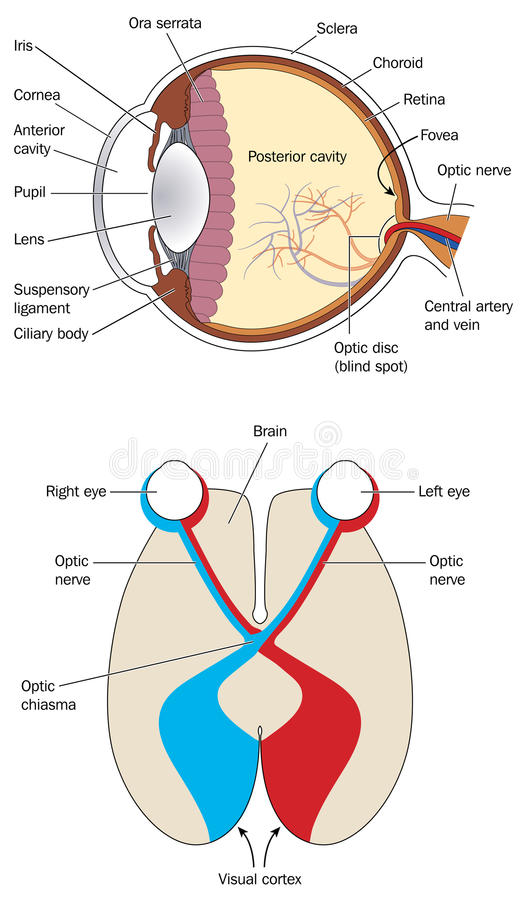
Chez l’homme, le cortex visuel est composé de six zones :
V1 – première analyse des informations
V2 – Vision stéréoscopique
V3 – distance et profondeur
V4 – couleur
V5 – mouvement
V6 – position objective d’un objet
Ensuite ces zones entrent en relation :
Circuit V1-V2-V3-V5-V6 : position de l’objet dans l’espace
Circuit V1-V2-V4 : nature de l’objet



La vision produite par œil-nerf optique-cerveau, un produit de la génétique, du développement et de l’évolution, vue… par des grands scientifiques, de Darwin à Gould en passant par Feynman
« Je me rappelle bien du temps où la pensée de l’œil me donnait froid partout, mais j’ai surmonté cette étape des rechignements... »
Lettre de Charles Darwin à Asa Gray daté du 3 avril, 1860
La conception de l’œil chez les animaux est reliée à l’évolution des espèces, au développement de l’individu et à la génétique
Le gène qui contrôle le développement de l’œil est le même chez tous les mammifères. Cela n’est pas surprenant. Les données fournies par les fossiles impliquent une branche commune pour tous les mammifères.
La découverte des gènes homéotiques qui pilotent la fabrication du corps a changé notre vision… de l’œil !
La génétique est apparue pilotée non seulement par des gènes mais par des structures d’interactions en série des gènes, avec des niveaux hiérarchiques de ces interventions. Un seul gène peut ainsi piloter, en cascade, jusqu’à 2500 gènes comme le gène maître de l’œil.
On s’est aperçu qu’un gène d’ours peut très bien fonctionner sur une fourmi et inversement. Si on inocule un gène qui commande la fabrication d’un oeil de mouche à une mouche drosophile, il lui pousse un oeil supplémentaire. Mais que se passe-t-il si on inocule un gène d’oeil de souris à cette mouche drosophile ? Le premier motif d’étonnement c’est que le gène de souris fonctionne très bien sur une mouche. Mais que va-t-il produire ? Est-ce un oeil de souris, un oeil de mouche ou une bizarrerie ? On pourrait se dire que cela devrait être un oeil de souris puisque le gène vient d’une souris ... En effet, on sait que la souris n’a pas du tout la même structure de l’oeil que la mouche. Eh bien non, c’est un oeil tout à fait normal de mouche qui va apparaître sur la drosophile ! Et l’inverse est vrai également : si on inocule à une grenouille un gène de fourmi, il poussera un oeil de plus et ce sera un oeil de grenouille. On a montré que la commande de fabrication d’un oeil en général est utilisable sur n’importe quel animal capable de faire fonctionner un oeil. On démontre ainsi que ce gène donne seulement l’ordre « fait pousser un oeil » et que cet ordre est commun aux diverses espèces vivantes, ou du moins interchangeable. Des gènes homéotiques, comme celui de l’oeil, sont ceux qui pilotent non seulement la formation d’un organe, mais tout le développement embryonnaire.
Walter Gehring, dans « La drosophile aux yeux rouges » :
« Le gène maître « eyeless » de la drosophile et celui de la souris sont capables d’induire la morphogenèse de l’œil. Plus récemment, nous avons aussi cloné le gène homologue d’eyeless chez le calmar, en collaboration avec Joram Piatigorsky et ses collaborateurs, et nous avons montré qu’il est aussi capable d’induire des yeux lorsqu’il est exprimé ectopiquement chez la drosophile. Puisque nous avons aussi observé des homologues de Pax-6 hautement conservés chez des ascidies, des vers némertiens et des vers plats, et montré que Pax-6 s’exprime spécifiquement dans les yeux, nous considérons qu’eyeless (Pax-6) est un gène maître universel contrôlant la morphogenèse de l’oeil. Ces résultats poussent, en outre, à conclure que l’œil prototypique n’est peut-être apparu qu’une seule fois, et non pas quarante fois, dans l’évolution. La grande diversité des types d’yeux que l’on trouve dans le règne animal aurait eu ce prototype pour point de départ des évolutions divergentes, parallèles ou convergentes ayant pris place ensuite. En fait, l’œil de certains vers plats ressemble étroitement au prototype darwinien et, chez une espèce trouvée au Japon, l’œil consiste en un seul photorécepteur et ue seule cellule pigmenaire, comme cela a été postulé par Darwin. »
François Jacob, dans son exposé de l’Université de tous les Savoirs, le 1er janvier 2000 :
« On voit les changements apportés dans la manière de considérer l’évolution biochimique. Tant que chaque gène, donc chaque protéine, était regardé comme un objet unique, résultat d’une séquence unique de nucléotides ou d’acides aminés, chacun d’eux ne pouvait se former que par une création nouvelle, de toute évidence hautement improbable. » François Jacob affirmait dans sa conférence pour l’Université de tous les savoirs : « On a longtemps pensé que les molécules de différents organismes étaient entièrement différentes. Et même que c’était la nature de leurs molécules qui donnait aux organismes leurs propriétés et particularités. En d’autres termes, que les chèvres avaient des molécules de chèvres et les escargots des molécules d’escargot. Que c’étaient les molécules de chèvre qui donnaient à la chèvre ses particularités. (...) Ce qui distingue un papillon d’un lion ou une poule d’une mouche, c’est moins une différence dans les constituants chimiques que dans l’organisation et la distribution de ces constituants. Par exemple, ce qui rend un vertébré différent d’un autre c’est plus un changement dans le temps d’expression et dans les quantités relatives des produits des gènes au cours du développement de l’embryon que les petites différences observées dans la structure de ces produits. (...) Chez la mouche, qui jouit d’un long passé génétique, ont été mis en évidence les gènes qui assurent, dans l’œuf, la mise en place des axes du futur embryon, puis ceux qui découpent le corps de l’embryon en segments puis ceux qui déterminent le destin et la forme de chacun de ces segments. A la stupéfaction générale, ces mêmes gènes ont été retrouvés chez tous les animaux examinés : coup sur coup grenouille, ver, souris et homme. Qui eut dit, il y a encore quinze ans, que les gènes qui mettent en place le plan d’un être humain sont les mêmes que ceux qui fonctionnent chez une mouche ou un ver ? (...) On voit les changements apportés dans la manière de considérer l’évolution biochimique. Tant que chaque gène, donc chaque protéine, était regardé comme un objet unique, résultat d’une séquence unique de nucléotides ou d’acides aminés, chacun d’eux ne pouvait se former que par une création nouvelle, de toute évidence hautement improbable. (...) Ce qui distingue un papillon d’un lion, une poule d’une mouche, c’est moins une différence dans les constituants chimiques que dans l’organisation et la distribution de ces constituants. Ce qui les rend différents c’est plus un changement dans le temps d’expression et dans la quantité relative des différents produits des gènes au cours du développement de l’embryon que les petites différences observées dans la structure de ces produits. »
Gerald Schroeder, dans "L’évolution, rationalité ou hasard ?" :
« Avec l’avènement de l’aptitude, par la biologie moléculaire, à discerner la structure des protéines et des gènes, la comparaison statistique de la similarité de ces structures parmi les animaux est devenue possible. Le gène qui contrôle le développement de l’œil est le même chez tous les mammifères. Cela n’est pas surprenant. Les données fournies par les fossiles impliquent une branche commune pour tous les mammifères.
Mais ce qui est surprenant, pour ne pas dire stupéfiant, c’est la similarité du gène de mammifère qui contrôle le développement des yeux chez les mollusques et dans les systèmes visuels chez les vers. On peut dire la même chose pour le gène qui contrôle le développement de membres chez les insectes et chez les humains. En fait, ce gène est si similaire que des morceaux du gène des mammifères, si on les introduit dans une mouche de fruit, feront apparaître une aile sur la mouche.
Cela aurait un sens si le développement de la vie était décrit comme un arbre. Mais le buisson de vie signifie que juste au-dessus du niveau de la vie unicellulaire, les insectes, les mammifères, les vers et les mollusques se sont séparés.
Le gène de l’œil a 130 sites. Cela veut dire qu’il y a 20130 (20 à la puissance 130, c’est-à-dire 20 multiplié 130 fois par lui-même) combinaisons possibles d’acides aminés le long de ces sites. Pour les raisons qui lui sont propres, la nature a sélectionné la même combinaison d’acides aminés pour tous les systèmes visuels de tous les animaux. Cette fidélité ne peut pas avoir été causée par hasard. Elle doit avoir été préprogrammée dans des formes inférieures de vie. Mais ces formes inférieures de vie, constituées par une seule cellule, n’avaient pas d’yeux.
Ces données ont déconcerté la théorie classique de " l’évolution aléatoire et indépendante " qui aurait produit ces structures convergentes. Cette similarité est si totalement insoupçonnée par les théories classiques de l’évolution, que la plus prestigieuse des revues scientifiques des Etats-Unis, Science, a rapporté : " L’hypothèse selon laquelle l’œil du céphalopode [mollusque] a évolué en convergence avec celui des vertébrés [humains] est remise en question par nos découvertes récentes du [gène] Pax-6... Le concept selon lequel les yeux des invertébrés ont évolué de manière complètement indépendante de l’œil des vertébrés devra être réexaminé. "
Il ne faut pas perdre de vue la portée de cette affirmation. On nous demande de réexaminer l’idée que l’évolution est un agent libre. La convergence, la similarité de ces gènes, est si grande qu’elle n’a pas pu arriver, qu’elle n’est pas arrivée par des réactions purement aléatoires. »
Darwin, dans « L’Origine des espèces » :
« La raison me dit que si l’on peut montrer qu’il existe de nombreuses étapes intermédiaires entre un œil imparfait et un œil complexe, parfait, chaque étape ayant son utilité pour l’individu… alors on pourra affirmer que la sélection naturelle est capable de produire un œil complexe, parfait, bien que cela soit difficile à imaginer, sans risquer de mettre en péril la rigueur interne de la théorie. »
« Pouvons-nous croire que la sélection naturelle puisse produire, d’une part, des organes insignifiants tels que la queue de la girafe, qui sert de chasse-mouches et, d’autre part, un organe aussi important que l’œil ? (…) Il semble absurde au possible, je le reconnais, de supposer que la sélection naturelle ait pu former l’œil avec toutes les inimitables dispositions qui permettent d’ajuster le foyer à diverses distances, d’admettre une quantité variable de lumière et de corriger les aberrations sphériques et chromatiques. Lorsqu’on affirma pour la première fois que le soleil est immobile et que la terre tourne autour de lui, le sens commun de l’humanité déclara la doctrine fausse ; mais on sait que le vieux dicton : Vox populi, vox Dei, n’est pas admis en matière de science. La raison nous dit que si, comme cela est certainement le cas, on peut démontrer qu’il existe de nombreuses gradations entre un œil simple et imparfait et un œil complexe et parfait, chacune de ces gradations étant avantageuse à l’être qui la possède ; que si, en outre, l’œil varie quelquefois et que ces variations sont transmissibles par hérédité, ce qui est également le cas ; que si, enfin, ces variations sont utiles à un animal dans les conditions changeantes de son existence, la difficulté d’admettre qu’un œil complexe et parfait a pu être produit par la sélection naturelle, bien qu’insurmontable pour notre imagination, n’attaque en rien notre théorie. Nous n’avons pas plus à nous occuper de savoir comment un nerf a pu devenir sensible à l’action de la lumière que nous n’avons à nous occuper de rechercher l’origine de la vie elle-même ; toutefois, comme il existe certains organismes inférieurs sensibles à la lumière, bien que l’on ne puisse découvrir chez eux aucune trace de nerf, il ne paraît pas impossible que certains éléments du sarcode, dont ils sont en grande partie formés, puissent s’agréger et se développer en nerfs doués de cette sensibilité spéciale. C’est exclusivement dans la ligne directe de ses ascendants que nous devons rechercher les gradations qui ont amené les perfectionnements d’un organe chez une espèce quelconque. Mais cela n’est presque jamais possible, et nous sommes forcés de nous adresser aux autres espèces et aux autres genres du même groupe, c’est-à-dire aux descendants collatéraux de la même souche, afin de voir quelles sont les gradations possibles dans les cas où, par hasard, quelques-unes de ces gradations se seraient transmises avec peu de modifications. En outre, l’état d’un même organe chez des classes différentes peut incidemment jeter quelque lumière sur les degrés qui l’ont amené à la perfection. L’organe le plus simple auquel on puisse donner le nom d’œil, consiste en un nerf optique, entouré de cellules de pigment, et recouvert d’une membrane transparente, mais sans lentille ni aucun autre corps réfringent. Nous pouvons, d’ailleurs, d’après M. Jourdain, descendre plus bas encore et nous trouvons alors des amas de cellules pigmentaires paraissant tenir lieu d’organe de la vue, mais ces cellules sont dépourvues de tout nerf et reposent simplement sur des tissus sarcodiques. Des organes aussi simples, incapables d’aucune vision distincte, ne peuvent servir qu’à distinguer entre la lumière et l’obscurité. Chez quelques astéries, certaines petites dépressions dans la couche de pigment qui entoure le nerf sont, d’après l’auteur que nous venons de citer, remplies de matières gélatineuses transparentes, surmontées d’une surface convexe ressemblant à la cornée des animaux supérieurs. M. Jourdain suppose que cette surface, sans pouvoir déterminer la formation d’une image, sert à concentrer les rayons lumineux et à en rendre la perception plus facile. Cette simple concentration de la lumière constitue le premier pas, mais de beaucoup le plus important, vers la constitution d’un œil véritable, susceptible de former des images ; il suffit alors, en effet, d’ajuster l’extrémité nue du nerf optique qui, chez quelques animaux inférieurs, est profondément enfouie dans le corps et qui, chez quelques autres, se trouve plus près de la surface, à une distance déterminée de l’appareil de concentration, pour que l’image se forme sur cette extrémité. Dans la grande classe des articulés, nous trouvons, comme point de départ, un nerf optique simplement recouvert d’un pigment ; ce dernier forme quelquefois une sorte de pupille, mais il n’y a ni lentille ni trace d’appareil optique. On sait actuellement que les nombreuses facettes qui, par leur réunion, constituent la cornée des grands yeux composés des insectes, sont de véritables lentilles, et que les cônes intérieurs renferment des filaments nerveux très singulièrement modifiés. Ces organes, d’ailleurs, sont tellement diversifiés chez les articulés, que Müller avait établi trois classes principales d’yeux composés, comprenant sept subdivisions et une quatrième classe d’yeux simples agrégés. Si l’on réfléchit à tous ces faits, trop peu détaillés ici, relatifs à l’immense variété de conformation qu’on remarque dans les yeux des animaux inférieurs ; si l’on se rappelle combien les formes actuellement vivantes sont peu nombreuses en comparaison de celles qui sont éteintes, il n’est plus aussi difficile d’admettre que la sélection naturelle ait pu transformer un appareil simple, consistant en un nerf optique recouvert d’un pigment et surmonté d’une membrane transparente, en un instrument optique aussi parfait que celui possédé par quelque membre que ce soit de la classe des articulés. Quiconque admet ce point ne peut hésiter à faire un pas de plus, et s’il trouve, après avoir lu ce volume, que la théorie de la descendance, avec les modifications qu’apporte la sélection naturelle, explique un grand nombre de faits autrement inexplicables, il doit admettre que la sélection naturelle a pu produire une conformation aussi parfaite que l’œil d’un aigle, bien que, dans ce cas, nous ne connaissions pas les divers états de transition. On a objecté que, pour que l’œil puisse se modifier tout en restant un instrument parfait, il faut qu’il soit le siège de plusieurs changements simultanés, fait que l’on considère comme irréalisable par la sélection naturelle. Mais, comme j’ai essayé de le démontrer dans mon ouvrage sur les variations des animaux domestiques, il n’est pas nécessaire de supposer que les modifications sont simultanées, à condition qu’elles soient très légères et très graduelles. Différentes sortes de modifications peuvent aussi tendre à un même but général ; ainsi, comme l’a fait remarquer M. Wallace, « si une lentille a un foyer trop court ou trop long, cette différence peut se corriger, soit par une modification de la courbe, soit par une modification de la densité ; si la courbe est irrégulière et que les rayons ne convergent pas vers un même point, toute amélioration dans la régularité de la courbe constitue un progrès. Ainsi, ni la contraction de l’iris, ni les mouvements musculaires de l’œil ne sont essentiels à la vision : ce sont uniquement des progrès qui ont pu s’ajouter et se perfectionner à toutes les époques de la construction de l’appareil. » Dans la plus haute division du règne animal, celle des vertébrés, nous pouvons partir d’un œil si simple, qu’il ne consiste, chez le branchiostome, qu’en un petit sac transparent, pourvu d’un nerf et plein de pigment, mais dépourvu de tout autre appareil. Chez les poissons et chez les reptiles, comme Owen l’a fait remarquer, « la série des gradations des structures dioptriques est considérable. » Un fait significatif, c’est que, même chez l’homme, selon Virchow, qui a une si grande autorité, la magnifique lentille cristalline se forme dans l’embryon par une accumulation de cellules épithéliales logées dans un repli de la peau qui affecte la forme d’un sac ; le corps vitré est formé par un tissu embryonnaire sous-cutané. Toutefois, pour en arriver à une juste conception relativement à la formation de l’œil avec tous ses merveilleux caractères, qui ne sont pas cependant encore absolument parfaits, il faut que la raison l’emporte sur l’imagination ; or, j’ai trop bien senti moi-même combien cela est difficile, pour être étonné que d’autres hésitent à étendre aussi loin le principe de la sélection naturelle. La comparaison entre l’œil et le télescope se présente naturellement à l’esprit. Nous savons que ce dernier instrument a été perfectionné par les efforts continus et prolongés des plus hautes intelligences humaines, et nous en concluons naturellement que l’œil a dû se former par un procédé analogue. Mais cette conclusion n’est-elle pas présomptueuse ? Avons-nous le droit de supposer que le Créateur met en jeu des forces intelligentes analogues à celles de l’homme ? Si nous voulons comparer l’œil à un instrument optique, nous devons imaginer une couche épaisse d’un tissu transparent, imbibé de liquide, en contact avec un nerf sensible à la lumière ; nous devons supposer ensuite que les différentes parties de cette couche changent constamment et lentement de densité, de façon à se séparer en zones, ayant une épaisseur et une densité différentes, inégalement distantes entre elles et changeant graduellement de forme à la surface. Nous devons supposer, en outre, qu’une force représentée par la sélection naturelle, ou la persistance du plus apte, est constamment à l’affût de toutes les légères modifications affectant les couches transparentes, pour conserver toutes celles qui, dans diverses circonstances, dans tous les sens et à tous les degrés, tendent à permettre la formation d’une image plus distincte. Nous devons supposer que chaque nouvel état de l’instrument se multiplie par millions, pour se conserver jusqu’à ce qu’il s’en produise un meilleur qui remplace et annule les précédents. Dans les corps vivants, la variation cause les modifications légères, la reproduction les multiplie presque à l’infini, et la sélection naturelle s’empare de chaque amélioration avec une sûreté infaillible. Admettons, enfin, que cette marche se continue pendant des millions d’années et s’applique pendant chacune à des millions d’individus ; ne pouvons-nous pas admettre alors qu’il ait pu se former ainsi un instrument optique vivant, aussi supérieur à un appareil de verre que les œuvres du Créateur sont supérieures à celles de l’homme ? (…)La sélection naturelle ne produit pas la perfection absolue ; autant que nous en pouvons juger, d’ailleurs, ce n’est pas à l’état de nature que nous rencontrons jamais ces hauts degrés. Selon Müller, la correction pour l’aberration de la lumière n’est pas parfaite, même dans le plus parfait de tous les organes, l’œil humain. Helmholtz, dont personne ne peut contester le jugement, après avoir décrit dans les termes les plus enthousiastes la merveilleuse puissance de l’œil humain, ajoute ces paroles remarquables : « Ce que nous avons découvert d’inexact et d’imparfait dans la machine optique et dans la production de l’image sur la rétine n’est rien comparativement aux bizarreries que nous avons rencontrées dans le domaine de la sensation. Il semblerait que la nature ait pris plaisir à accumuler les contradictions pour enlever tout fondement à la théorie d’une harmonie préexistante entre les mondes intérieurs et extérieurs. (…)L’organe le plus simple auquel on puisse donner le nom d’œil, consiste en un nerf optique, entouré de cellules de pigment, et recouvert d’une membrane transparente, mais sans lentille ni aucun autre corps réfringent. Nous pouvons, d’ailleurs, d’après M. Jourdain, descendre plus bas encore et nous trouvons alors des amas de cellules pigmentaires paraissant tenir lieu d’organe de la vue, mais ces cellules sont dépourvues de tout nerf et reposent simplement sur des tissus sarcodiques. Des organes aussi simples, incapables d’aucune vision distincte, ne peuvent servir qu’à distinguer entre la lumière et l’obscurité. Chez quelques astéries, certaines petites dépressions dans la couche de pigment qui entoure le nerf sont, d’après l’auteur que nous venons de citer, remplies de matières gélatineuses transparentes, surmontées d’une surface convexe ressemblant à la cornée des animaux supérieurs. M. Jourdain suppose que cette surface, sans pouvoir déterminer la formation d’une image, sert à concentrer les rayons lumineux et à en rendre la perception plus facile. Cette simple concentration de la lumière constitue le premier pas, mais de beaucoup le plus important, vers la constitution d’un œil véritable, susceptible de former des images ; il suffit alors, en effet, d’ajuster l’extrémité nue du nerf optique qui, chez quelques animaux inférieurs, est profondément enfouie dans le corps et qui, chez quelques autres, se trouve plus près de la surface, à une distance déterminée de l’appareil de concentration, pour que l’image se forme sur cette extrémité.
Dans la grande classe des articulés, nous trouvons, comme point de départ, un nerf optique simplement recouvert d’un pigment ; ce dernier forme quelquefois une sorte de pupille, mais il n’y a ni lentille ni trace d’appareil optique. On sait actuellement que les nombreuses facettes qui, par leur réunion, constituent la cornée des grands yeux composés des insectes, sont de véritables lentilles, et que les cônes intérieurs renferment des filaments nerveux très singulièrement modifiés. Ces organes, d’ailleurs, sont tellement diversifiés chez les articulés, que Müller avait établi trois classes principales d’yeux composés, comprenant sept subdivisions et une quatrième classe d’yeux simples agrégés.
Si l’on réfléchit à tous ces faits, trop peu détaillés ici, relatifs à l’immense variété de conformation qu’on remarque dans les yeux des animaux inférieurs ; si l’on se rappelle combien les formes actuellement vivantes sont peu nombreuses en comparaison de celles qui sont éteintes, il n’est plus aussi difficile d’admettre que la sélection naturelle ait pu transformer un appareil simple, consistant en un nerf optique recouvert d’un pigment et surmonté d’une membrane transparente, en un instrument optique aussi parfait que celui possédé par quelque membre que ce soit de la classe des articulés.
Quiconque admet ce point ne peut hésiter à faire un pas de plus, et s’il trouve, après avoir lu ce volume, que la théorie de la descendance, avec les modifications qu’apporte la sélection naturelle, explique un grand nombre de faits autrement inexplicables, il doit admettre que la sélection naturelle a pu produire une conformation aussi parfaite que l’œil d’un aigle, bien que, dans ce cas, nous ne connaissions pas les divers états de transition. On a objecté que, pour que l’œil puisse se modifier tout en restant un instrument parfait, il faut qu’il soit le siège de plusieurs changements simultanés, fait que l’on considère comme irréalisable par la sélection naturelle. Mais, comme j’ai essayé de le démontrer dans mon ouvrage sur les variations des animaux domestiques, il n’est pas nécessaire de supposer que les modifications sont simultanées, à condition qu’elles soient très légères et très graduelles. Différentes sortes de modifications peuvent aussi tendre à un même but général ; ainsi, comme l’a fait remarquer M. Wallace, « si une lentille a un foyer trop court ou trop long, cette différence peut se corriger, soit par une modification de la courbe, soit par une modification de la densité ; si la courbe est irrégulière et que les rayons ne convergent pas vers un même point, toute amélioration dans la régularité de la courbe constitue un progrès. Ainsi, ni la contraction de l’iris, ni les mouvements musculaires de l’œil ne sont essentiels à la vision : ce sont uniquement des progrès qui ont pu s’ajouter et se perfectionner à toutes les époques de la construction de l’appareil. » Dans la plus haute division du règne animal, celle des vertébrés, nous pouvons partir d’un œil si simple, qu’il ne consiste, chez le branchiostome, qu’en un petit sac transparent, pourvu d’un nerf et plein de pigment, mais dépourvu de tout autre appareil. Chez les poissons et chez les reptiles, comme Owen l’a fait remarquer, « la série des gradations des structures dioptriques est considérable. » Un fait significatif, c’est que, même chez l’homme, selon Virchow, qui a une si grande autorité, la magnifique lentille cristalline se forme dans l’embryon par une accumulation de cellules épithéliales logées dans un repli de la peau qui affecte la forme d’un sac ; le corps vitré est formé par un tissu embryonnaire sous-cutané. Toutefois, pour en arriver à une juste conception relativement à la formation de l’œil avec tous ses merveilleux caractères, qui ne sont pas cependant encore absolument parfaits, il faut que la raison l’emporte sur l’imagination ; or, j’ai trop bien senti moi-même combien cela est difficile, pour être étonné que d’autres hésitent à étendre aussi loin le principe de la sélection naturelle.
La comparaison entre l’œil et le télescope se présente naturellement à l’esprit. Nous savons que ce dernier instrument a été perfectionné par les efforts continus et prolongés des plus hautes intelligences humaines, et nous en concluons naturellement que l’œil a dû se former par un procédé analogue. Mais cette conclusion n’est-elle pas présomptueuse ? Avons-nous le droit de supposer que le Créateur met en jeu des forces intelligentes analogues à celles de l’homme ? Si nous voulons comparer l’œil à un instrument optique, nous devons imaginer une couche épaisse d’un tissu transparent, imbibé de liquide, en contact avec un nerf sensible à la lumière ; nous devons supposer ensuite que les différentes parties de cette couche changent constamment et lentement de densité, de façon à se séparer en zones, ayant une épaisseur et une densité différentes, inégalement distantes entre elles et changeant graduellement de forme à la surface. Nous devons supposer, en outre, qu’une force représentée par la sélection naturelle, ou la persistance du plus apte, est constamment à l’affût de toutes les légères modifications affectant les couches transparentes, pour conserver toutes celles qui, dans diverses circonstances, dans tous les sens et à tous les degrés, tendent à permettre la formation d’une image plus distincte. Nous devons supposer que chaque nouvel état de l’instrument se multiplie par millions, pour se conserver jusqu’à ce qu’il s’en produise un meilleur qui remplace et annule les précédents. Dans les corps vivants, la variation cause les modifications légères, la reproduction les multiplie presque à l’infini, et la sélection naturelle s’empare de chaque amélioration avec une sûreté infaillible. Admettons, enfin, que cette marche se continue pendant des millions d’années et s’applique pendant chacune à des millions d’individus ; ne pouvons-nous pas admettre alors qu’il ait pu se former ainsi un instrument optique vivant, aussi supérieur à un appareil de verre que les œuvres du Créateur sont supérieures à celles de l’homme ? »
Stephen Jay Gould, dans « La structure de la théorie de l’évolution » :
« On est passé de la préférence pour un adaptationnisme point par point (chaque organe étant expliqué comme le résultat, relativement indépendant de toute contrainte, d’un façonnement distinct par la sélection naturelle en raison d’une utilité présente) à la reconnaissance que des voies développementales homologues (conservées en tant qu’héritage d’un lointain passé, dans un cadre original) façonnent puissamment « depuis l’intérieur » les possibilités actuelles. Ce changement général dans la façon de voir s’est étendu aux études phylogénétiques de tous niveaux, depuis l’examen de certaines similitudes entre les phyla les plus disparates jusqu’à l’analyse de certaines variations morphologiques entre espèces appartenant à de petits rameaux monophylétiques au sein de l’arbre phylogénétique du règne animal. Aucun autre cas n’a fait l’objet de plus d’attention, engendré plus de surprises, reposé sur de plus solides observations et modifié autant les certitudes antérieures que la découverte d’une importante voie développementale, clairement homologue, sous-tendant un phénomène qui était encore, il n’y pas si longtemps, dans les manuels un vénérable exemple type de convergence : il s’agit de l’apparition évolutive indépendante, chez plusieurs phyla, d’yeux à cristallin, capables de former des images, les similitudes anatomiques stupéfiantes des yeux dotés d’un cristallin unique chez les céphalopodes et chez les vertébrés en étant l’illustration la plus frappante. Comme Tomarev et al. (1997) l’écrivent : « On avait considéré jusqu’ici que les yeux complexes des mollusques céphalopodes et ceux des vertébrés représentaient l’exemple classique d’une évolution convergente. » Il reste légitime de parler de convergence anatomique pour le domaine limité des résultats finals ; mais, en ce qui concerne la voie développementale par laquelle ces structures sont édifiées, c’est maintenant le phénomène de signification théorique opposée qui tient le haut du pavé… Salvini-Plawen et Mayr (1977) ont soutenu que des photorécepteurs d’une forme ou d’une autre sont apparus au cours de l’évolution environ 40 à 60 fois indépendamment dans le règne animal, six phyla ayant élaboré des yeux complexes capables de former des images. Les six phyla en question sont… les cnidaires, les annélides, les onychophores, les arthropodes, les mollusques et les vertébrés. Au début des années 1990, en utilisant des sondes de Drosphila, les chercheurs clonèrent une famille de gènes mamaliens appelés « Pax », dont notamment le gène Pax-6. Celui-ci est partculièrement remarquable, comprenant à la fois une boîte « paired » et une homéoboîte (Walter et Gruss, 1991). Peu de temps parès, on a rapporté à des altérations de Pax-6 plusieurs mutations déjà connues, affectant la forme ou la fonction des yeux. Par exemple, les souris hétérozygotes pour le gène muté small eye (Sey) présentent des yeux de dimension réduite, tandis que les homozygotes, qui ne sont pas viables, ne développent pas d’yeux du tout. Les recherches ont ensuite montré que Pax-6 s’exprime dans la moelle épinière, ainsi que dans plusieurs régions du cerveau, et plus particulièrement lors de la morphogénèse des yeux chez les vertébrés… (Gehring, 1996)… Peu de chercheurs avaient imaginé que la version présente chez Drosophila fonctionnerait aussi de la même manière fondamentale… La fonction similaire chez des phyla différents des gènes homéotiques de l’œil a été démontrée de façon spectaculaire en faisant s’exprimer le gène de souris chez Drosophila (Halder et al., 1995) : la version mammalienne s’est montrée capable d’induire l’édification d’yeux surnuméraires. Voilà qui confirmait la qualification de « gène régulateur maître » du développement des yeux donnée par Gehring à Pax-6… Mais la découverte de cette conservation évolutive de la voie développementale sous-tendant l’édification des yeux d’insectes et de vertébrés, si surprenante qu’elle soit, étant donné les suppositions antérieures sur l’impossibilité de telles homologies entre gènes chez des phyla éloignés, ne permettait pas encore d’examiner directement le problème théorique de la convergence dans l’évolution. Tout compte fait, l’œil à cristallin unique des vertébrés n’a que peu de similitude anatomique avec l’œil de mouche à multiples facettes, et personne n’a jamais proclamé à leur sujet qu’ils témoignaient d’un phénomène de convergence. Mais le fait qu’une voie développementale homologue sous-tende l’édification d’yeux morphologiquement disparates chez deux phyla très différents soulevait une question évidente : pouvait-on considérer Pax-6 comme un « gène régulateur maître » (pour reprendre de nouveau l’expression de Gehring) contrôlant le développement de tous les yeux complexes, y compris ceux à cristallin unique des vertébrés et des céphalopodes, dont les remarquables similitudes de fonction et de structure en font le « paradigme de l’évolution convergente » (Gehring, 1996) ? (…)
Tomarev et al. (1997) ont réalisé une observation très attendue : ils ont trouvé un homologue du gène Pax-6 des vertébrés et des arthropodes chez le calmar Loligo opalescens. Chez ce céphalopode, ce gène s’exprime dans les yeux embryonnaires, dans les organes olfactifs, le cerveau et les tentacules… Ces chercheurs ont aussi obtenu un résultat particulièrement satisfaisant : l’expression ectopique du gène Pax-6 du calmar induit également la formation d’yeux surnuméraires chez Drosophila…
Gehring (1996, 1998) fonde l’expression qu’il a proposée, « gène régulateur maître », sur trois caractéristiques de Pax-6 : c’est le régulateur amont de toute une importante série de gènes déterminant plus spécifiquement l’édification des yeux ; il est interchangeable entre les phyla, tout en restant toujours le déclencheur d’une série aval d’actions conduisant à l’édification des yeux « propres » à chaque animal donné ; et il est capable d’une façon générale de susciter l’édification d’yeux surnuméraires en des localisations inhabituelles…
Les découvertes concernant Pax-6 ont maintenant montré l’existence d’une importante base homologique entre les voies de développement sous-tendant l’édification des yeux complexes des céphalopodes et des vertébrés. Ainsi, un canal, constitué par une contrainte interne héritée, a énormément facilité la réalisation d’une solution quasi identique chez les deux phyla, et les évolutionnistes ne peuvent plus continuer à soutenir que des yeux aussi similaires sont apparus évolutivement selon deux voies entièrement distinctes, sous la seule action de la sélection naturelle, et sans l’aide d’aucun canal commun représenté par un même processus d’édification au cours du développement. Mais, tout comme les partisans de la pure convergence se trompaient en affirmant que le mécanisme qu’ils invoquaient était intervenu en exclusivité, la découverte des homologies de Pax-6 ne permet pas de passer complètement à une explication invoquant exclusivement la contrainte.
Comme c’est fréquemment le cas, étant donné l’organisation hiérarchique du monde organique, une certaine explication (par exemple, la convergence) est valable à un niveau, et une explication alternative (par exemple, la contrainte) à un autre. »
Jay Gould, dans son essai « The Problem of Perfection » :
« La sélection naturelle joue un rôle constructif dans le système de Darwin. Il échafaude progressivement l’adaptation, au moyen d’une séquence de stades intermédiaires, et réunit, de manière séquentielle, les éléments qui semblent souvent avoir de sens seulement en tant qu’éléments d’un produit final. Mais comment une série de formes intermédiaires raisonnables peut-elle être construite ? Quel peut être la valeur, pour l’organisme qui en est le possesseur, de la première étape minuscule vers un œil ? »
« Le principe général avancé par les évolutionnistes modernes pour résoudre ce dilemme fait appel à un concept portant le nom malheureux de “ préadaptation ”. (Je dis bien malheureux, car le terme implique que les espèces s’adaptent à l’avance, à des événements imminents dans leur évolution historique, tandis que c’est plutôt le sens contraire exactement qui est recherché.) (...) Nous évitons la question excellente : à quoi sert cinq pour cent d’un œil en faisant valoir que le possesseur d’une telle structure naissante ne l’ait pas utilisé pour la vue. »
Richard Feynman, Cours de Physique – Mécanique tome 2 – :
« Le phénomène des couleurs dépend partiellement du monde physique. Par exemple, les couleurs dans les pellicules de savon sont produites par des interférences. Mais cela dépend aussi, bien sûr, de l’œil et de ce qui se passe derrière l’œil dans le cerveau.
La physique permet de caractériser la lumière qui pénètre dans l’œil, mais après cela nos sensations sont les résultats de processus neuro-photochimiques et de réponses psychologiques.
On trouve de nombreux phénomènes intéressants associés avec la vision qui comportent un mélange de phénomènes physiques et de processus physiologiques, et la pleine compréhension des phénomènes naturels, comme nous les « voyons », doit dépasser la physique au sens habituel. Nous ne nous excusons pas de faire ces excursions dans d’autres domaines, parce que la séparation des domaines ne relève que de la commodité humaine, et n’est pas une chose naturelle. La nature n’est pas intéressée par nos séparations et nombreux sont les phénomènes intéressants qui sont à cheval sur divers domaines…
Nous étudions ici un domaine dans lequel la physique et d’autres sciences sont très étroitement liées. Ce domaine, c’est la « vision ». En particulier nous allons examiner la « vision en couleur »…
Tout commence avec l’œil ; aussi, afin de comprendre quels phénomènes nous voyons, nous devons un peu savoir ce qu’est l’œil. Au chapitre suivant nous envisagerons dans les détails comment les diverses parties de l’œil fonctionnent et comment elles sont reliées avec le système nerveux. Pour le présent, nous ne décrirons que brièvement comment l’œil fonctionne.
La lumière pénètre dans l’œil au travers de la « cornée » ; elle est déviée et vient former une image sur une couche appelée la « rétine » à l’arrière de l’œil, de telle sorte que les différentes parties de la rétine reçoivent la lumière des différentes parties du champ visuel externe. La rétine n’est pas absolument uniforme : il y a un endroit, une tache au centre de notre champ de vision que nous utilisons lorsque nous essayons de voir les choses avec beaucoup de soin et pour lequel nous avons la plus grande acuité de vision. Cet endroit est appelé la « tache jaune » ou « macula ».
Comme nous pouvons immédiatement nous en rendre compte à partir de l’expérience en observant des objets, les parties latérales de l’œil ne permettent pas une vision aussi détaillée que le centre. Il existe également un endroit sur la rétine d’où partent les nerfs transportant toute l’information ; c’est un point aveugle. Il n’y a pas de région sensible de la rétine à cet endroit, et il est possible de démontrer que si nous fermons par exemple l’œil gauche, que nous regardions directement quelque chose, nous fermons par exemple l’œil gauche, que nous regardions directement quelque chose, et qu’ensuite nous déplaçions un doigt ou un autre petit objet lentement hors du champ de vision, il disparaisse soudainement quelque part…
En différents endroits de la rétine, il y a différents types de structures. Les objets qui apparaissent avec plus de densité près de la périphérie de la rétine sont appelés des « bâtonnets ». Plus près de la tache jaune, nous trouvons à côté de ces cellules à bâtonnet, des cellules en « cônes »…
Lorsque nous nous rapprochons de la tache jaune, le nombre de cônes augmente et dans la tache jaune elle-même il n’y a rien d’autre en fait que des cellules en cône, étroitement serrées, si étroitement que les cellules en cône sont beaucoup plus fines et beaucoup plus étroites à cet endroit que n’importe où ailleurs.
Nous devons donc comprendre que nous voyons au milieu du champ de vision grâce aux cônes, mais que pour les bords nous utilisons les autres cellules, les bâtonnets. Il est maintenant intéressant de remarquer que dans la rétine chaque cellule qui est sensible à la lumière n’est pas directement reliée par une fibre au nerf optique, mais est reliée à beaucoup d’autres cellules qui elles-mêmes sont reliées les unes aux autres.
On trouve différents types de cellules : il y a les cellules qui transportent l’information vers le nerf optique, mais il y a les autres qui sont essentiellement reliées « horizontalement ». On trouve essentiellement quatre types de cellules, mais nous n’entrerons pas dans ces détails maintenant.
La chose importante sur laquelle nous insistons est que le signal de la lumière a déjà subi un premier « traitement réfléchi ». C’est-à-dire que l’information venant des diverses cellules ne va pas immédiatement vers le cerveau, endroit par endroit, mais que dans la rétine cette quantité d’information a déjà été digérée par une combinaison de l’information venant de plusieurs récepteurs visuels. Il est important de comprendre que certains phénomènes du type de ceux qui se passent dans le cerveau apparaissent dans l’œil lui-même.
Un des phénomènes les plus frappants de la vision est celui de l’adaptation de l’œil à l’obscurité. Si nous pénétrons dans l’obscurité sortant d’une chambre brillamment éclairée, il ne nous est pas possible de très bien voir pendant un certain temps et puis, petit à petit, les choses deviennent de plus en plus apparentes et éventuellement nous pouvons discerner quelque chose là où précédemment nous ne voyions rien. Si l’intensité de la lumière est très basse, les objets que nous voyons n’ont « pas de couleurs ». Il est reconnu que cette vision adaptée à l’obscurité est presque entièrement due aux bâtonnets, tandis que la vision à la lumière brillante est due aux cônes. Comme résultat, un certain nombre de phénomènes peuvent aisément se comprendre à cause du transfert de fonctions des cônes et des bâtonnets ensemble aux seuls bâtonnets.
On trouve de nombreux cas pour lesquels si l’intensité de la lumière avait été plus forte, nous aurions pu voir la couleur et nous trouverions ces choses magnifiques. Par exemple, au travers d’un télescope nous voyons presque toujours en « noir et blanc » des images de nébuleuses faibles, mais W. C. Miller des Observatoires du Mont Wilson et de Palomar a eu la patience de faire des photographies « en couleur » de certains de ces objets. Personne n’a en réalité vu ces couleurs avec son œil, mais ce ne sont pas des couleurs artificielles, cela est dû simplement au fait que l’intensité de la lumière n’est pas suffisamment grande pour que les cônes de notre œil puissent les voir…
En lumière brillante, les bâtonnets ont en apparence une très faible sensibilité mais, dans l’obscurité, lorsque le temps passe, ils récupèrent alors leur capacité de voir la lumière. Les variations de l’intensité de la lumière pour lesquelles on peut s’adapter dépassent le rapport de un million à un. La nature ne réalise pas tout cela avec un seul type de cellule, mais elle fait effectuer son travail tantôt par des cellules capables de voir la lumière brillante – les cellules voyant en couleur, les cônes – tantôt par des cellules de basse intensité adaptées à l’obscurité, les bâtonnets. Parmi les conséquences intéressantes de ce transfert, on trouve en premier lieu que, à cette occasion, les couleurs disparaissent et deuxièmement qu’il y a une différence dans la luminosité relative des objets colorés différemment. Il se trouve que les bâtonnets voient mieux dans le bleu que ne le font les cônes et que les cônes peuvent voir par exemple une lumière d’un rouge profond tandis que les bâtonnets se trouvent dans l’incapacité totale de la voir. Ainsi la lumière rouge est noire en qui concerne les bâtonnets…
Un autre effet provenant du fait que les bâtonnets deviennent prééminents dans le noir et du fait de l’absence de bâtonnets dans la tache jaune, est que lorsque nous observons directement quelque chose dans l’obscurité, notre vision n’est pas aussi précise que lorsque nous le regardons de côté…
Un autre effet intéressant dû au fait que le nombre de cônes diminue lorsque nous nous éloignons sur le côté du champ de vision est que, même en lumière brillante, la couleur disparaît lorsque l’objet s’éloigne de côté. La manière de le vérifier consiste à regarder dans une certaine direction fixe, à demander à un ami de se rapprocher latéralement en tenant des cartes colorées et d’essayer de préciser de quelles couleurs elles sont avant qu’elles ne soient directement en face de vous. On trouve qu’on peut déterminer que les cartes sont bien là bien avant que l’on puisse en déterminer la couleur. Quand on fait cela, il vaut mieux venir du côté opposé au point aveugle, sinon c’est assez troublant de percevoir la couleur, puis de ne rien voir, puis de la revoir.
Un autre phénomène intéressant est que la périphérie de la rétine est très sensible au mouvement. Bien que nous ne puissions voir très correctement depuis le coin de notre œil, si un quelconque insecte se déplace et que nous ne nous attendions pas à ce que quelque chose d’autre se déplace à cet endroit, nous y sommes immédiatement sensibles. Nous sommes tous « fabriqués » de telle sorte que nous puissions voir quelque chose s’agiter sur le côté de notre champ de vision.
Nous nous intéressons maintenant à la vision des cônes, à la vision la plus brillante et nous en venons à la question qui est la plus caractéristique de la vision par les cônes, c’est-à-dire la couleur. Comme nous le savons, la lumière blanche peut être décomposée par un prisme en tout un spectre de longueurs d’onde qui nous apparaissent comme ayant des couleurs différentes ; c’est exactement ce que sont les couleurs, bien entendu : des apparences. N’importe quelle source de lumière peut être analysée par un réseau ou un prisme et on peut déterminer la distribution spectrale, c’est-à-dire la « quantité » de chaque longueur d’onde. Une certaine lumière peut avoir beaucoup de bleu, un rouge en quantité, très peu de jaune, etc. Tout cela est extrêmement précis du point de vue de la physique, mais la question est de savoir de quelle « couleur » cela apparaîtra-t-il ? Il est évident que les différentes couleurs dépendent d’une manière ou d’une autre de la distribution spectrale de la lumière, mais le problème est de trouver quelles caractéristiques de la distribution spectrale produisent les diverses sensations. Par exemple, que devons-nous faire pour obtenir une couleur verte ? Nous savons tous que nous pouvons prendre simplement une partie du spectre qui est verte. Mais est-ce le « seul » moyen d’obtenir du vert ou de l’orange, ou n’importe quelle autre couleur ?
Y a-t-il plus d’une distribution spectrale qui produise le même effet visuel apparent ? La réponse est certainement « oui ». Il y a un nombre très limité d’effets visuels, en fait une multiplicité à trois dimensions comme nous allons le voir très bientôt, mais on peut tracer un nombre infini de courbes différentes pour représenter la lumière provenant de différentes sources. La question maintenant que nous devons discuter est : sous quelles conditions des contributions différentes de la lumière apparaissent à l’œil comme étant exactement de la même couleur ?
La technique psychologique la plus puissante dans le jugement des couleurs consiste à utiliser l’œil comme un « instrument de zéro ». C’est-à-dire : nous n’essayons pas de définir ce qui constitue une sensation verte, parce qu’il se trouve que ceci est extrêmement compliqué. Au lieu de cela, nous étudions les conditions dans lesquelles deux stimulis « ne peuvent être distingués ». Alors il n’est pas nécessaire de décider si deux personnes ont la même sensation dans des circonstances différentes, mais seulement si lorsque pour une personne deux sensations sont les mêmes, elles le restent pour une autre. Nous n’avons pas à dire si, lorsque quelqu’un voit du vert, ce qu’il ressent est la même chose que ce que ressent quelqu’un d’autre lorsqu’il voit quelque chose de vert ; de cela nous ne savons rien.
Pour illustrer ces possibilités, nous pouvons utiliser une série de quatre lampes projecteurs, montées avec des filtres et dont les luminosités sont ajustables continûment dans un grand domaine : l’une dispose d’un filtre rouge et envoie une tache rouge sur l’écran, la suivante dispose d’un filtre vert et forme une tache verte, la troisième a un filtre bleu et la quatrième est un cercle blanc avec une tache noire en son centre. Si maintenant nous mettons un petit peu de lumière rouge et qu’à côté d’elle nous mettions un petit peu de vert, nous voyons que sur la surface de recouvrement cela produit une sensation qui n’est pas ce que nous appelons un vert rougeâtre, mais une nouvelle couleur, le jaune dans ce cas particulier. En modifiant les proportions de rouge et de vert, nous pouvons passer par diverses teintes d’orange, etc…
Nous ne pouvons certainement pas obtenir toutes les différentes couleurs en ne mélangeant que le rouge et le vert, parce que, par exemple, le bleu n’apparaît jamais dans un tel mélange. Cependant, en ajoutant un peu de bleu dans la région centrale où les trois taches se chevauchent, on peut obtenir quelque chose qui apparaît comme un très joli blanc….
Voici une des grandes lois de la couleur : si deux distributions spectrales ne peuvent être distinguées et que nous ajoutions à chacune d’elles une certaine lumière, les nouveaux mélanges ne peuvent également pas être distingués…
Le second principe de mélange des couleurs de lumières est celui-ci : n’importe quelle couleur peut être formée à partir de trois couleurs différentes, par exemple rouge, verte et bleue…
Notre question est de savoir pourquoi les couleurs se comportent-elles ainsi ? La théorie la plus simple proposée par Young et Helmoltz, suppose que dans l’œil se trouvent différents pigments qui reçoivent la lumière et qu’ils ont différents spectres d’absorption, de telle sorte qu’un pigment absorbe fortement par exemple dans le rouge, un autre absorbe fortement dans le bleu et un autre absorbe dans le vert… Les pigments qui peuvent être extraits d’une rétine sont formés essentiellement d’un pigment appelé « pourpre visuel ». Les caractéristiques les plus remarquables de celui-ci sont, premièrement, qu’il se trouve dans l’œil de pratiquement tous les vertébrés, et, deuxièmement, que sa courbe de réponse s’adapte magnifiquement à la sensibilité de l’œil… Ce pigment est manifestement le pigment avec lequel nous voyons dans l’obscurité : le pourpre visuel est le pigment des bâtonnets, et il n’a rien à voir avec la vision en couleur… C’est Rushton qui a détecté les pigments de couleur de l’œil… Il utilise un ophtalmoscope, envoyant de la lumière dans l’œil au travers du cristallin, puis focalisant la lumière qui revient, et mesurant la quantité de lumière qui est réfléchie… Les cônes sont conçus de manière que la lumière qui entre dans le cône rebondit dans tous les sens et se fraye un chemin jusqu’aux petits points sensibles, aux sommets. La lumière descend directement vers le point sensible, rebondit au fond et ressort en ayant traversé une quantité considérable de pigment de la vision en couleur…
Les premières théories de la vision disaient qu’il y a trois pigments et trois types de cônes, chacun contenant un pigment ; qu’un nerf part de chaque cône vers le cerveau de telle sorte que les trois parcelles d’information sont transportées au cerveau ; et puis que tout se passe dans le cerveau. Ceci, bien sûr, est une idée incomplète : il ne sert à rien de découvrir que l’information est transportée le long du nerf optique au cerveau, parce que nous n’avons même pas commencé de résoudre le problème. Nous devons nous poser des questions plus fondamentales. L’endroit où les informations sont rassemblées a-t-il une importance ? Est-il important qu’elles soient transportées directement dans le cerveau par le nerf optique ou bien la rétine peut-elle faire d’abord un peu de mise en ordre ? Nous avons vu une représentation de la rétine où il apparaissait que c’était une chose extrêmement compliquée avec beaucoup de connexions internes et où pouvaient déjà débuter quelques analyses. »
En fait, les gens qui étudient l’anatomie et le développement de l’œil ont montré que la rétine est, en réalité, dans le cerveau : dans le développement de l’embryon, un morceau de cerveau sort vers l’avant et de longues fibres poussent vers l’arrière reliant les yeux au cerveau. La rétine est organisée exactement de la même manière que le cerveau et, comme quelqu’un l’a dit d’une manière très jolie, « Le cerveau a développé un moyen qui lui permet de regarder le monde ». L’œil est une partie du cerveau qui rencontre pour ainsi dire la lumière. Ainsi, il n’est pas du tout improbable qu’une partie de l’analyse de la couleur ait déjà été analysée dans la rétine.
Ceci nous donne une possibilité extrêmement intéressante. Aucun des autres sens comporte une aussi grande quantité de calculs, pour ainsi dire, avant que le signal pénètre dans un nerf sur lequel on puisse faire des mesures. Les calculs pour tous les autres sens se passent d’habitude dans le cerveau, et il est très difficile de se rendre en des endroits particuliers du cerveau pour y faire des mesures parce qu’il y a trop d’interconnexions. Ici, avec la sensation visuelle, nous avons la lumière, trois couches de cellules opérant les calculs, et les résultats de ces calculs sont transmis par le nerf optique. Ainsi nous avons la première occasion d’observer physiologiquement comment, peut-être, les premières couches du cerveau fonctionnent dans leurs premières étapes. Cela présente donc un double intérêt, n’intéressant pas seulement la vision, mais intéressant l’ensemble de la physiologie.
Le fait qu’il y ait trois pigments ne signifie pas qu’il doive y avoir trois types de sensations. Une des autres théories de la vision en couleur dit qu’il y a en réalité des systèmes de couleurs opposées. c’est-à-dire qu’une des fibres nerveuses transporte beaucoup d’impulsions si on voit du jaune, et moins que d’habitude pour le bleu. Une autre fibre nerveuse transporte des informations vert et rouge de la même manière, et une autre le blanc et le noir. En d’autres termes, dans cette théorie quelqu’un a déjà commencé de faire une hypothèse sur le système de connexions et sur la méthode de calcul…
La rétine est, en fait, semblable à la surface du cerveau… Toute partie de la rétine est reliée à d’autres parties et l’information qui en sort portée sur les longs axones qui forment le nerf optique est des combinaisons de l’information venue de nombreuses cellules. Il y a trois couches de cellules avec une succession de fonctions : il y a les cellules de la rétine, qui sont celles que la lumière impressionne, une cellule intermédiaire qui prend l’information venant d’une seule ou de quelques cellules de la rétine et qui la transfère à nouveau à diverses cellules d’une troisième couche et qui la transporte au cerveau. Il y a toutes sortes de liaisons croisées entre les cellules d’une même couche…
La focalisation de la lumière est accomplie essentiellement par la cornée, par le fait qu’elle a une surface courbe qui « dévie » la lumière. C’est pourquoi nous ne pouvons voir clairement sous l’eau, parce qu’alors la différence entre l’indice de la cornée, qui est 1,37, et celui de l’eau qui est 1,33 est insuffisante. Derrière la cornée, il y a de l’eau, avec pratiquement un indice de 1,33 et derrière encore se trouve une lentille de structure extrêmement intéressante : c’est une série de couches empilées comme dans un oignon, à ceci près qu’elle est entièrement transparente et qu’elle a un indice 1,40 au centre et 1,38 à l’extérieur… De plus, la forme de la cornée n’est pas celle d’une sphère. Une lentille sphérique est toujours affligée d’aberration sphérique. La cornée est plus « plate » à l’extérieur que ne l’est une sphère, de telle sorte que les aberrations sphériques sont plus faibles pour la cornée qu’elles ne le seraient pour une lentille sphérique placée en cet endroit. La lumière est focalisée sur la rétine par le système lentille-cornée. Lorsque nous regardons des objets qui sont plus ou moins éloignés, la lentille se resserre ou se détend et modifie le foyer pour s’adapter aux distances différentes. Pour ajuster la quantité totale de lumière, il y a l’iris qui est ce que nous appelons la couleur de l’œil, un œil brun ou bleu, selon la personne, lorsque la quantité de lumière augmente et diminue, l’iris se referme et s’écarte.
Considérons maintenant le mécanisme nerveux pour contrôler l’accommodation de la lentille, le mouvement de l’œil, les muscles qui tournent l’œil dans sa cavité et l’iris. De toutes les informations qui sortent du nerf optique, la grande majorité divisée en deux paquets est ensuite envoyée au cerveau. Mais il y a quelques fibres, qui vont nous intéresser maintenant, qui ne vont pas directement au cortex visuel du cerveau où nous « voyons » les images, mais qui vont dans le cerveau intermédiaire. Ce sont les fibres qui mesurent la lumière moyenne et réalisent l’ajustement de l’iris ; ou si l’image paraît floue, elles essayent de corriger la lentille, ou s’il y a une double image elles essayent d’ajuster l’œil à la vision binoculaire. En tous les cas, elles vont vers le cerveau intermédiaire et reviennent dans l’œil. Il y a des muscles qui commandent l’accommodation de la lentille et un autre qui pénètre dans l’iris. L’iris dispose de deux systèmes musculaires. L’un est un muscle circulaire qui, lorsqu’il est excité, se contracte et ferme l’iris ; il agit très rapidement et les nerfs sont directement reliés au cerveau par le biais d’axones courts allant jusque dans l’iris. Les muscles opposés sont des muscles radials, de telle sorte que lorsque l’extérieur devient sombre et que le muscle circulaire se détend, ces muscles radials tirent dans l’autre sens. Nous avons ici, comme dans de nombreux endroits dans le corps, un ensemble de muscles qui fonctionnent dans des directions opposées et dans chacun de ces cas, le système nerveux qui contrôle les deux est délicatement ajusté, de telle sorte que lorsque des signaux sont envoyés afin de raidir le premier, des signaux sont automatiquement envoyés pour détendre l’autre. L’iris est une exception particulière : les nerfs qui font se contracter l’iris sortent d’un endroit que personne ne connaît exactement, descendent dans la moelle dans le dos, atteignent les sections thoraciques, sortent de la moelle, passent par les ganglions du cou et retournent vers la tête afin de faire fonctionner l’autre extrémité de l’iris.
En fait, le signal passe au travers d’un système nerveux complètement différent, qui n’est pas du tout le système nerveux central, mais le système nerveux sympathique, c’est donc une manière étrange de faire fonctionner le système.
Nous avons déjà insisté sur une autre chose étrange à propos de l’œil, qui est que les cellules sensibles à la lumière sont du mauvais côté, de sorte que la lumière doit traverser de nombreuses couches d’autres cellules avant qu’elle n’atteigne ces récepteurs – elle est construite avec le dedans à l’extérieur ! Ainsi certaines caractéristiques sont splendides et d’autres sont en apparence stupides…
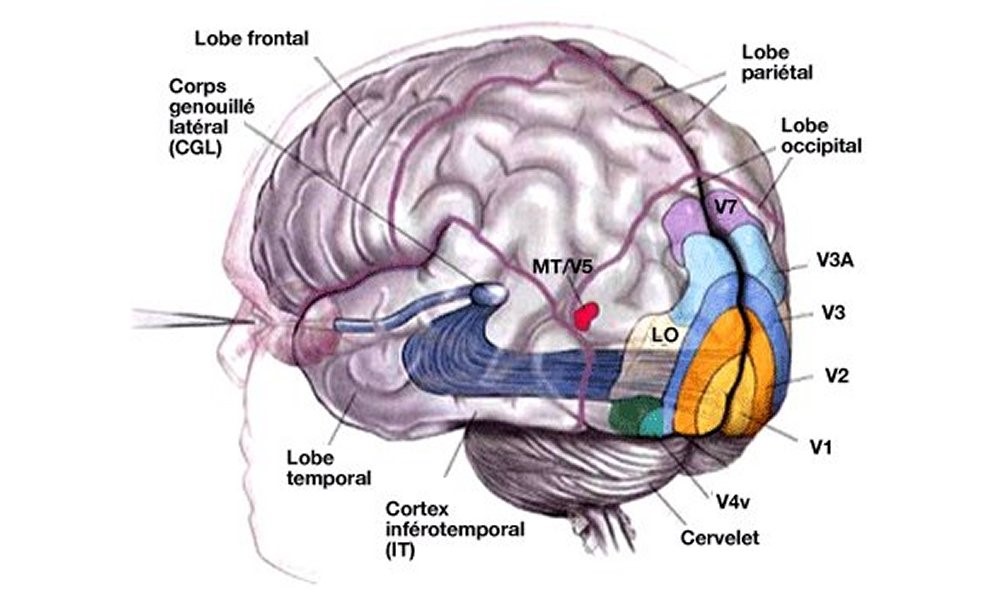
Les fibres du nerf optique pénètrent dans une certaine région du cerveau, juste au-dessus du génicule latéral, d’où elles sortent vers une section du cerveau appelée le cortex visuel. Remarquez que certaines des fibres venant de chaque œil sont envoyées de l’autre côté du cerveau… Les nerfs optiques du côté gauche de l’œil droit passent au travers du chiasme optique, tandis que ceux du côté gauche de l’œil gauche passent à côté et se déplacent de la même manière. Ainsi la partie gauche du cerveau reçoit toutes les informations qui viennent du côté gauche des globes de chaque œil, c’est-à-dire concernant le côté droit du champ visuel, tandis que le côté droit du cerveau voit le côté gauche du champ visuel. C’est ainsi que l’information de chacun des deux yeux est rassemblée afin de pouvoir dire à quelle distance se trouvent les objets. C’est le système de vision binoculaire.
Les connexions entre la rétine et le cortex visuel sont intéressantes. Si une zone de la rétine est enlevée ou détruite d’une manière ou d’une autre, alors toute la fibre va mourir, et nous pouvons donc trouver avec quelle zone elle est reliée. Il apparaît que les connexions sont avant tout biunivoques – chaque région de la rétine correspond à une région dans le cortex visuel – et les régions qui sont très proches sur la rétine sont très proches dans le cortex visuel. Ainsi le cortex visuel reproduit encore la disposition spatiale des bâtonnets et des cônes, mais, bien entendu, très déformée. Les images qui sont au centre du champ, qui occupent une très petite partie de la rétine, sont étalées sur de très nombreuses cellules dans le cortex visuel…
D’une manière ou d’une autre, l’information doit aller d’un côté du cerveau à l’autre par l’intermédiaire d’autres canaux, ce qui est tout à fait surprenant. La question de savoir comment ce réseau s’est « établi » est également très intéressante. Le problème de savoir quelle partie est déjà formée et quelle partie est « apprise » est un vieux problème… Un autre phénomène intéressant est lié au mouvement de l’œil. Les yeux doivent se déplacer afin de faire se coïncider les deux images dans divers cas. Ces mouvements sont de types différents : l’un consiste à suivre quelque chose, ce qui nécessite que les deux yeux doivent aller dans la même direction, à droite ou à gauche, et l’autre est de les pointer au même endroit à différentes distances, ce qui nécessite qu’ils doivent se déplacer en sens opposé.
Les nerfs pénétrant dans les muscles des yeux sont déjà structurés pour de tels buts. Il y a un ensemble de nerfs qui tirent les muscles à l’intérieur d’un œil et à l’extérieur de l’autre, et relâchent les muscles opposés, de telle sorte que les deux yeux se déplacent ensemble. Il existe un autre centre où une excitation fait s’écarter les yeux de la parallèle et les fait se déplacer l’un vers l’autre. Chaque œil peut tourner vers l’extérieur lorsque l’autre œil se déplace vers le nez, mais il est impossible consciemment ou inconsciemment de tourner en même temps les deux yeux vers l’extérieur non pas parce qu’il n’y a pas de muscles, mais parce qu’il n’y a aucune manière d’envoyer un signal qui fasse tourner les deux yeux vers l’extérieur, sauf si nous avons subi un accident ou qu’il y ait quelque chose d’anormal, par exemple un nerf qui ait été coupé... Nous sommes déjà structurés à un certain degré. C’est un point important, parce que la plupart des premiers livres sur l’anatomie et la psychologie, etc., ne réalisaient pas ou n’insistaient pas sur le fait que nous sommes déjà si complètement structurés – ils disaient que tout n’est qu’apprentissage…
L’œil humain n’est pas le seul type d’œil. Dans les vertébrés, presque tous les yeux sont essentiellement semblables à l’œil humain. Cependant, chez les animaux inférieurs, il y a beaucoup d’autres types d’yeux : des yeux en taches, divers yeux en coupes et d’autres yeux moins sensibles, que nous n’avons pas le temps de décrire. Mais il y a un autre œil extrêmement développé chez les invertébrés, l’œil « composé » de l’insecte…
On a découvert ainsi que l’œil de l’abeille est sensible sur un plus grand domaine spectral que le nôtre. Notre œil fonctionne entre 7.000 Angströms et 4.000 Angströms, du rouge au violet, mais l’abeille peut voir jusqu’à 3.000 Angströms dans l’ultra-violet ! Ceci rend compte d’un grand nombre d’effets différents et intéressants. En premier lieu, les abeilles peuvent distinguer de nombreuses fleurs qui nous paraissent semblables. Bien sûr, nous devons réaliser que les couleurs des fleurs ne sont pas conçues pour « notre » œil, mais pour celui de l’abeille, ce sont des signaux pour attirer les abeilles vers une fleur particulière… Un autre aspect intéressant de la vision de l’abeille est que l’abeille peut donner la direction du soleil en regardant un coin de ciel bleu sans voir le soleil lui-même. Nous ne pouvons aisément le faire. Lorsque nous regardons le ciel par le fenêtre et que nous voyons qu’il est bleu, dans quelle direction se trouve le soleil ? L’abeille peut le savoir, parce que l’abeille est sensible à la polarisation de la lumière et la lumière diffusée du ciel est polarisée…
On dit également que l’abeille peut remarquer des battements d’ailes jusqu’à 200 périodes par seconde, tandis que nous en voyons au maximum jusqu’à 20. Le mouvement des abeilles dans les ruches est très rapide ; les pieds se déplacent, les ailes vibrent, mais il nous est très difficile de distinguer ces mouvements avec notre œil…
L’œil d’une abeille est un œil composé, et il est constitué d’un grand nombre de cellules spéciales appelées « ommatidia », qui sont disposées en cône sur la surface d’une sphère (approximativement) à l’extérieur de la tête de l’abeille… Au sommet il y a une surface transparente, une sorte de « lentille », mais en réalité cela ressemble plus à un filtre ou à un guide de lumière qui fait descendre la lumière vers la fibre étroite qui est l’endroit où probablement se produit l’absorption. A l’autre extrémité de celle-ci sort la fibre nerveuse. La fibre centrale est entourée sur ses côtés par six cellules qui, en fait, ont sécrété la fibre…
Les plus grands yeux au monde sont ceux du calamar géant ; on en a trouvé avec un diamètre allant jusqu’à 40 cm !
Un des aspects de notre sujet est celui des interconnexions de l’information entre une partie de l’œil et une autre. Considérons l’œil composé du crabe des moluques sur lequel on a fait beaucoup d’expériences. D’abord, nous devons connaître le type d’information qui peut se déplacer le long des nerfs. Un nerf transporte un type de perturbation qui a un effet électrique facile à détecter, un type de perturbation de caractère ondulatoire qui parcourt le nerf et produit un effet à l’autre extrémité : un long morceau de cette cellule nerveuse, appelé l’axone, transporte l’information, et un certain type d’impulsion appelée « excitation » se déplace le long de l’axone s’il est excité à une extrémité. Lorsqu’une excitation parcourt le nerf, une autre ne peut immédiatement suivre. Toutes les excitations ont la même taille, ainsi nous n’obtenons pas d’excitations plus intenses lorsque le nerf est plus fortement excité, mais nous obtenons davantage d’excitations par seconde. La dimension de l’excitation est déterminée par la fibre. Il est important d’avoir compris cela afin de voir ce qui se passe ensuite.
L’œil du crabe des molluques ne ressemble pas beaucoup à un œil, et il est simplement constitué d’un millier d’ommatidia… Si nous envoyons de la lumière à un certain instant et mesurons les impulsions électriques qui se produisent, nous observons un léger retard au départ, et puis une série rapide de décharges qui graduellement se ralentit jusqu’à une vitesse uniforme. Lorsque la lumière cesse, les décharges s’arrêtent. Il est très intéressant de remarquer que si, alors que notre amplificateur est toujours relié à cette même fibre nerveuse, nous envoyons la lumière sur un ommatidium différent, rien ne se passe ; pas de signal.
Nous réalisons maintenant une autre expérience : nous envoyons de la lumière sur l’ommatidium original et obtenons la même réponse, et si maintenant nous envoyons en même temps la lumière sur un ommatidium voisin, les impulsions s’interrompent brièvement et réapparaissent ensuite à une vitesse beaucoup plus lente. Le taux de l’un est limité par les impulsions qui sortent de l’autre ommatidium ! En d’autres termes, chaque fibre nerveuse transporte l’information venant de l’ommatidium mais la quantité qu’elle transporte est limitée par les signaux venant des autres. Ainsi, par exemple, si l’œil dans son ensemble est plus ou moins uniformément éclairé, l’information venant de chacune des ommatidia sera relativement faible parce qu’elle est limitée par un grand nombre d’autres. En fait la limitation est additive – si nous envoyons la lumière sur plusieurs ommatidia voisines, la limitation devient très grande. La limitation est plus grande lorsque les ommatidia sont plus proches et si les ommatidia sont suffisamment éloignées l’une de l’autre, la limitation est pratiquement nulle. Elle est donc additive et dépend de la distance ; voilà un premier exemple de la combinaison dans l’œil même de l’information venant de différentes parties de l’œil. Nous pouvons voir, par exemple, en y réfléchissant un peu, que ceci est un procédé pour « augmenter le contraste » aux bords des objets, parce que si une partie de la scène est lumineuse et une autre partie sombre, les ommatidia dans la surface éclairée envoient des impulsions qui sont limitées par la lumière dans le voisinage, et l’effet sera relativement faible. D’un autre côté, un ommatidium à la frontière qui reçoit une impulsion « blanche » est également limité par ses voisins, mais il y en a moins, puisque certains sont dans l’obscurité ; le signal résultant est en conséquence plus élevé… Le crabe verra un renforcement des contours.
L’existence d’un renforcement des contours est connue depuis longtemps ; en fait, c’est une chose remarquable qui a été souvent commentée par les psychologues. Pour dessiner un objet, il nous suffit de dessiner son contour ! Qu’est-ce que le contour ? Le contour est simplement la différence au bord entre le clair et le sombre ou une couleur et une autre. Ce n’est pas le fait, croyez-le ou pas, que chaque objet possède une ligne qui l’entoure. Une telle ligne n’existe pas. Cette ligne n’existe que dans notre propre construction psychologique ; nous commençons à comprendre les raisons pour lesquelles la « ligne » est suffisante pour obtenir l’objet total. Notre propre œil travaille probablement d’une manière semblable – beaucoup plus compliquée mais semblable.
Finalement, nous décrirons brièvement le travail très minutieux, le travail superbe et d’avant-garde qui a été réalisé sur la grenouille. En réalisant une expérience correspondante sur la grenouille, en plaçant des électrodes semblables à des aiguilles, très joliment construites, très fines, dans le nerf optique d’une grenouille, on peut obtenir les signaux qui se déplacent le long d’un axone particulier et exactement comme dans le cas du crabe des moluques, nous trouvons que l’information ne dépend pas que d’un seul endroit dans l’œil mais est une somme d’informations venant de différents endroits.
Le schéma le plus récent du fonctionnement de l’œil de la grenouille est le suivant. On peut trouver quatre différents types de fibres nerveuses optiques, en ce sens qu’il y a quatre différents types de réponses. Ces expériences n’ont pas été réalisées en envoyant des impulsions de lumière, parce que ce n’est pas ce que voit la grenouille. Une grenouille se trouve quelque part et son œil ne se déplace jamais, sauf lorsque la feuille de nénuphar sur laquelle elle se trouve se met à se balancer, et dans ce cas son œil oscille juste ce qu’il faut pour que l’image reste fixe. Elle ne tourne pas son œil. Si quelque chose se déplace dans son champ de vision, par exemple une petite mouche (elle doit être capable de voir quelque chose de petit se déplaçant dans un environnement fixe) quatre types différents de fibres envoient des décharges – leurs propriétés sont résumées ainsi :
Types de réponses des fibres du nerf optique d’une grenouille
1- Détection de bord soutenue (non effaçable) – vitesse : O,2-0,5 m/s – champ angulaire : 1°
2- Détection de bord convexe (effaçable) – vitesse : 0,5 m/s – champ angulaire : 2°-3°
3- Détection de changement de contraste – vitesse : 1-2 m/s – champ angulaire : 7 à 10°
4- Détection d’assombrissement – vitesse : jusqu’à ½ m/s – champ angulaire : jusqu’à 15°
5- Détection d’obscurité : très grand champ angulaire
Détection soutenue de bord, non effaçable, signifie que si nous introduisons dans le champ de vision de la grenouille un objet avec un bord, une grande quantité d’impulsions apparaissent dans cette fibre particulière tant que l’objet se déplace, mais elles diminuent jusqu’à former une suite continue d’impulsions qui se perpétue aussi longtemps que le bord est là, même s’il ne se déplace pas. Si nous éteignons la lumière, les impulsions s’arrêtent. Si nous l’allumons à nouveau, alors le bord est toujours en vue, elles réapparaissent. Elles ne sont pas effaçables. Un autre type de fibre lui ressemble beaucoup, à ceci près que si le bord est droit elle ne fonctionne pas. Ce doit être un bord convexe avec de l’obscurité derrière lui ! Quelle doit être la complexité du système d’interconnexion dans la rétine de l’œil de la grenouille pour qu’elle puisse comprendre qu’une surface convexe est entrée dans le champ de vision ? De plus, bien que cette fibre fonctionne assez longtemps, elle ne fonctionne pas aussi longtemps que l’autre et si nous éteignons la lumière puis la rallumons les impulsions ne reviennent pas. Cela dépend du mouvement d’arrivée de la surface convexe. L’œil la voit arriver et se souvient qu’elle est là, mais si nous arrêtons la lumière pour un instant, il l’oublie et ne la voit plus.
Un autre exemple est la détection de changement de contraste. Si un bord pénètre ou sort, il y a des impulsions apparaissent, mais si elle reste basse ou se tient élevée, les impulsions s’arrêtent ; cela ne fonctionne que lorsque la lumière diminue.
Finalement, il y a quelques fibres qui sont des détecteurs d’obscurité – une chose tout à fait étonnante – elles fonctionnent tout le temps ! Si nous augmentons la lumière, elles fonctionnent moins rapidement, mais tout le temps ! Si nous diminuons la lumière, elles fonctionnent plus rapidement, toujours tout le temps. Dans le noir, elles fonctionnent à toute allure, répétant perpétuellement : « Il fait noir ! Il fait noir ! Il fait noir ! »
Ces réponses semblent plutôt compliquées à classer et nous pouvons nous demander si les expériences ont peut-être été mal interprétées. Mais il est très intéressant de retrouver ces mêmes catégories clairement séparées dans l’anatomie de la grenouille ! Par d’autres mesures, après que ces réponses aient été classées (« après », c’est ce qui est important à ce propos), on a découvert que la vitesse des signaux sur les différentes fibres n’était pas la même, ce qui fait qu’il y a une autre manière indépendante de vérifier à quel type de fibre on a affaire ! (…)
Le tectum de la grenouille est là où les nerfs vont dans le cerveau en venant du nerf optique. Toutes les fibres nerveuses venant du nerf optique établissent des liaisons dans les diverses couches du tectum. Cette structure en couche est analogue à la rétine ; c’est en partie la raison pour laquelle nous savons que le cerveau et la rétine se ressemblent beaucoup. En prenant une électrode et en la déplaçant vers le bas au travers des différentes couches, nous pouvons trouver à quel endroit se termine un certain nerf optique et le beau et merveilleux résultat que l’on trouve est que les différents types de fibres se terminent dans différentes couches ! La première se termine dans la couche numéro un, la seconde au numéro deux, la troisième et cinquième se terminent au même endroit, et la plus profonde de toutes est la numéro quatre…
Il y a probablement trois pigments. Il peut y avoir différentes sortes de cellules réceptrices contenant les trois pigments dans des proportions différentes, mais on trouve beaucoup de connexions croisées qui permettent des additions et des soustractions par l’intermédiaire de l’addition et du réenforcement dans le système nerveux. Ainsi, avant de pouvoir vraiment comprendre la vision en couleur, il nous faudra comprendre la sensation finale. »

Oeil de grenouille